

Механизм обмена газов в легких и тканях. транспорт кислорода и углекислого газа.
Газообмен между альвеолярным воздухом и притекающей к легким венозной кровью — это совокупность процессов, обеспечивающих переход кислорода внешней среды в кровь, а углекислого газа из крови в альвеолы. Перемещение газов (легкие — кровь) осуществляется под влиянием разности парциальных давлений и напряжений этих газов в каждой из сред организма (табл.8.1).
Таблица 8.1.
Содержание и парциальное давление (напряжение) кислорода и углекислого газа в различных средах
| Среда | Кислород | Углекислый газ | ||||
| % | мм рт. ст. | мл/л | % | мм рт. ст. | мл/л | |
| Вдыхаемый воздух | 20,93 | 209,3 | 0,03 | 0,2 | 0,3 | |
| Выдыхаемый воздух | 16,0 | 160,0 | 4,5 | |||
| Альвеолярный воздух | 14,0 | 140,0 | 5,5 | |||
| Артериальная кровь | — | 100-96 | 200,0 | — | 560-540 | |
| Венозная кровь | — | 140-160 | — | |||
| Ткань | — | 10-15 | — | — | — | |
| Около митохондрий | — | 01-1 | — | — | — |
Альвеолярный воздух осуществляет газообмен с притекающей к легким венозной кровью, являясь как бы внутренней газовой средой организма. Состав альвеолярного воздуха отличается постоянством, мало изменяясь при обычном дыхании. При спокойном дыхании в альвеолы с каждым вдохом взрослого человека поступает 350 мл воздуха, и альвеолярный воздух обновляется лишь на 1/7 своего объема (коэффициент вентиляции). При спокойном дыхании давление в альвеолах ниже атмосферного.
Решающим фактором, обусловливающим непрерывность газообмена, является постоянство газового состава альвеолярного воздуха.
Учитывая свойство газов диффундировать из области большего парциального давления в область с меньшим парциальным давлением, несложно понять направленность диффузии О2 и СО2 на том или ином уровне дыхания (рис.8.6).
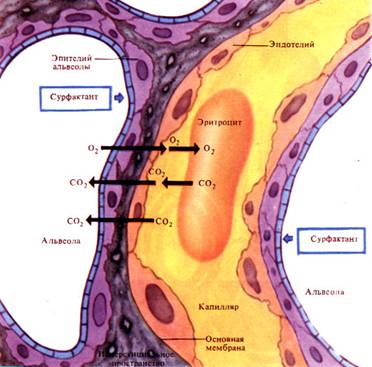
Рис.8.6. Газообмен в легких.
Парциальное давление кислорода в воздухе, заполняющем альвеолы легких, около 106 мм рт. ст., а его напряжение в плазме венозной крови, притекающей к легким, около 40 мм рт.ст. Вследствие разности давлений кислород из альвеол направляется в плазму крови и далее в эритроциты, где его напряжение практически равно нулю. Там он связывается с гемоглобином эритроцитов.
Парциальное давление углекислого газа в альвеолярном воздухе составляет 40 мм рт.ст., а его напряжение в притекающей к легким венозной крови — 46 мм рт.ст. Вследствие разности давлений углекислый газ переходит в альвеолы.
В артериальной крови, притекающей к тканям, напряжение кислорода выше, чем в тканях, а напряжение углекислого газа наоборот значительно ниже. Оно составляет 60 мм рт.ст. в ткани и 40 мм рт.ст. в плазме крови. В эритроцитах напряжение углекислого газа практически равно нулю. Вследствие этого кислород переходит из крови в ткани и включается в цикл метаболических процессов, а углекислый газ, в избытке содержащийся в тканях, переходит в кровь и переносится затем в легкие.
Процесс газообмена происходит непрерывно до тех пор, пока существует разность парциальных давлений и напряжений газов в каждой из сред, участвующих в газообмене.
Величина газообмена является показателем интенсивности окислительных процессов, протекающих в тканях. Для оценки интенсивности газообмена определяют количество кислорода, использованного организмом за определенное время, и количество углекислого газа, выделенного организмом за это же время. Об уровне газообмена можно судить и по величине минутной вентиляции легких. При спокойном дыхании через легкие проходит около 8000 мл воздуха в 1 мин. При физических или эмоциональных напряжениях, различных заболеваниях, сопровождающихся усилением окислительных процессов в тканях, легочная вентиляция возрастает. Газообмен между тканями и кровью, кровью и легкими, легкими и внешней средой может в значительной степени нарушаться при различных заболеваниях легких, сердечно-сосудистой системы, крови. Следствием таких нарушений газообмена может явиться гипоксия — кислородное голодание тканей.
Газообмен в организме осуществляется двумя основными механизмами:
1. Конвективный, представляет собой механическое передвижение молекул О2 и СО2 с током воздуха или крови. Таким образом, осуществляется перенос газов в воздухе или крови на большое расстояние.
2. Диффузия. Механизм газообмена между разными средами организма. Диффузия осуществляется из области с высоким парциальным давлением газов в область низкого их давления, причём на работу по переносу молекул затрачивается их собственная кинетическая энергия
В организме кислород и углекислый газ транспортируются кровью.
Кислород, поступающий из альвеолярного воздуха в кровь, связывается с гемоглобином эритроцитов, образуя так называемый оксигемоглобин, и в таком виде доставляется к тканям.
Количество кислорода, связанного гемоглобином в 100 мл крови, носит название кислородная ёмкость крови.
Известно, что каждый грамм гемоглобина связывает 1,34-1,35 мл О2. Следовательно, КЕК здорового мужчины, у которого в 100 мл крови содержится 15 г Hb, составляет 20,4 объёмных процента (табл.8.2).
Таблица 8.2. Количество гемоглобина и кислородная емкость крови
| Гемоглобин у мужчин у женщин | 14-15 гр% Нb (на 100 мл крови) 13,5-14,5 гр% Нb (на 100 мл крови) |
| Кислородная ёмкость крови КЕК) в покое при мышечной работе | 20 об % О2 (1 гр. Нb связывает 1,34-1,35 мл О2) увеличение на 5-10 % |
В тканевых капиллярах кислород отщепляется и переходит в ткани, где включается в окислительные процессы. Свободный гемоглобин связывает водород и превращается в так называемый восстановленный гемоглобин. Углекислый газ, образующийся в тканях, переходит в кровь и поступает в эритроциты. Затем часть углекислого газа соединяется с восстановленным гемоглобином, образуя так называемый карбогемоглобин, и в таком виде углекислый газ и доставляется к легким. Однако большая часть углекислого газа в эритроцитах при участии фермента карбоангидразы превращается в бикарбонаты, которые переходят в плазму и транспортируются к легким. В легочных капиллярах бикарбонаты при помощи специального фермента карбоангидразы распадаются и выделяется углекислый газ. Отщепляется углекислый газ и от гемоглобина. Углекислый газ переходит в альвеолярный воздух и с выдыхаемым воздухом удаляется во внешнюю среду.
Следует знать, что более эффективно, чем углекислый газ с гемоглобином, связывается окись углерода известная как угарный газ. Образующийся в этом случае так называемый карбоксигемоглобин не способен связывать кислород.
Наконец, последним этапом дыхания является тканевое дыхание или окислительно-восстановительные реакции, протекающие в клетках организма. Существо этих реакций заключается в том, что сложные органические вещества окисляются при участии специальных ферментов кислородом до конечных продуктов в виде аммиака, воды и двуокиси углерода. Освобождающаяся при этом энергия выделяется частично в виде тепла, однако основная ее часть идет на образование известных всем молекул АТФ, которые являются источником энергии, необходимой для жизнедеятельности организма.
В тканевых капиллярах кислород отщепляется и переходит в ткани, где включается в окислительные процессы. Свободный гемоглобин связывает водород и превращается в так называемый восстановленный гемоглобин.
§
Главным физиологическим стимулом дыхательных центров является двуокись углерода. Регуляция дыхания обусловливает поддержание нормального содержания СО2 в альвеолярном воздухе и артериальной крови. Возрастание содержания СО2 в альвеолярном воздухе на 0,17% вызывает удвоение минутного объёма дыхания, а вот снижение О2 на 39-40% не вызывает существенных изменений минутного объёма дыхания.
При повышении в замкнутых герметических кабинах концентрации СО2 до 5 — 8% у обследуемых наблюдалось увеличение легочной вентиляции в 7-8 раз. При этом концентрация СО2 в альвеолярном воздухе существенно не возрастала, так как основным признаком регуляции дыхания является необходимость регуляции объема легочной вентиляции, поддерживающей постоянство состава альвеолярного воздуха.
Деятельность дыхательного центра зависит от состава крови, поступающей в мозг по общим сонным артериям. В 1890 г. это было показано Фредериком в опытах с перекрестным кровообращением. У двух собак, находившихся под наркозом, перерезали и соединяли перекрестно сонные артерии и яремные вены. При этом голова первой собаки снабжалась кровью второй собаки и наоборот. Если у одной из собак, например у первой, перекрывали трахею и таким путем вызывали асфиксию, то гиперпноэ развивалось у второй собаки. У первой же собаки, несмотря на увеличение в артериальной крови напряжения СО2 и снижение напряжения 02, развивалось апноэ, так как в ее сонную артерию поступала кровь второй собаки, у которой в результате гипервентиляции снижалось напряжение СО2 в артериальной крови.
Двуокись углерода, водородные ионы и умеренная гипоксия вызывают усиление дыхания. Эти факторы усиливают деятельность дыхательного центра, оказывая влияние на периферические (артериальные) и центральные (модулярные) хеморецепторы, регулирующие дыхание.
Снижение РаО2 ниже нормы (- РаО2) получило название гипоксического стимула, повышение РаСО2 против нормы ( РаСО2) названо гиперкапническим стимулом,а снижение рН получило название ацидотического стимула.
Для дыхательного центра главным стимулом является — гиперкапнический. Реакция на гипоксию примерно в 4 раза меньше.
Артериальные хеморецепторы находятся в каротидных синусах и дуге аорты. Они расположены в специальных тельцах, обильно снабжаемых артериальной кровью. Аортальные хеморецепторы на дыхание влияют слабо, и большее значение имеют для регуляции кровообращения.
Артериальные хеморецепторы являются уникальными рецепторными образованиями, на которые гипоксия оказывает стимулирующее влияние. Афферентные влияния каротидных телец усиливаются также при повышении в артериальной крови напряжения двуокиси углерода и концентрации водородных ионов. Стимулирующее действие гипоксии и гиперкапнии на хеморецепторы взаимно усиливается, тогда, как в условиях гипероксии чувствительность хеморецепторов к двуокиси углерода резко снижается. Артериальные хеморецепторы информируют дыхательный центр о напряжении 02 и СО2 в крови, направляющейся к мозгу.
После перерезки артериальных (периферических) хеморецепторов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию и ацидоз.
Центральные хеморецепторы расположены в продолговатом мозге латеральнее пирамид. Перфузия этой области мозга раствором со сниженным рН резко усиливает дыхание, а при высоком рН дыхание ослабевает, вплоть до апноэ. То же происходит при охлаждении или обработке этой поверхности продолговатого мозга анестетиками. Центральные хеморецепторы, оказывая сильное влияние на деятельность дыхательного центра, существенно изменяют вентиляцию легких. Установлено, что снижение рН спинномозговой жидкости всего на 0,01 сопровождается увеличением легочной вентиляции на 4 л/мин.
Центральные хеморецепторы реагируют на изменение напряжения СО2 в артериальной крови позже, чем периферические хеморецепторы, так как для диффузии СО2 из крови в спинномозговую жидкость и далее в ткань мозга необходимо больше времени. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз — тормозят центральные хеморецепторы.
Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга, изучения синергизма и антагонизма дыхательных газов, взаимодействия системы дыхания и сердечно-сосудистой системы используют метод возвратного дыхания. При дыхании в замкнутой системе выдыхаемый СО2 вызывает линейное увеличение концентрации СО2 и одновременно повышается концентрация водородных ионов в крови, а также во внеклеточной жидкости мозга.
Совокупность дыхательных нейронов следовало бы рассматривать как созвездие структур, осуществляющих центральный механизм дыхания. Таким образом, вместо термина «дыхательный центр» правильнее говорить о системе центральной регуляции дыхания, которая включает в себя структуры коры головного мозга, определенные зоны и ядра промежуточного, среднего, продолговатого мозга, варолиева моста, нейроны шейного и грудного отделов спинного мозга, центральные и периферические хеморецепторы, а также механорецепторы органов дыхания.
§
| ЧСС, уд/мин | % МПК |
| 120-130 190 и выше | 50 % 60 % 70 % 75-80 % 90 % 100 % |
Разность между потребностью в кислороде и его потреблением составляет энергию, получаемую в результате анаэробного распада, и называется КИСЛОРОДНОМ ДОЛГОМ.
После окончания работы потребление кислорода остается еще некоторое время несколько более высоким по сравнению с уровнем покоя и медленно возвращается к этому уровню. Принято говорить, что в это время происходит оплата кислородного долга.
Кислородный долг состоит из двух компонентов или фракций — лактацидного и алактатного.
ЛАКТАЦИДНЫЙ ДОЛГ — представляет собой добавочную энергию, затрачиваемую на превращение главного побочного продукта анаэробного метаболизма — молочной кислоты (лактата) в пировиноградную кислоту.
АЛАКТАТНЫЙ ДОЛГ — представляет собой дополнительную энергию, необходимую для рефосфорилирования креатинфосфата и для восстановления запасов кислорода в мышечном миоглобине.
Общий расход энергии и факторы его определяющие при различных видах трудовой и спортивной деятельности
Суточный расход энергии у здорового человека значительно превышает величину основного обмена и складывается из следующих компонентов:
— основного обмена;
— рабочей прибавки, т. е. энергозатрат, связанных с движением и с выполнением той или иной работы;
— специфического-динамического действия пищи — увеличения интенсивности обмена веществ и энергозатрат, связанных с приемом пищи, процессами пищеварения и всасывания. Так прием белковой пищи увеличивает обмен на 30-40%, а при питании жирами и углеводами обмен увеличивается на 4-15%.
Энерготраты при физической работе резко увеличиваются. Например, при ходьбе расходуется энергии на 80—100% больше по сравнению с покоем, при беге — на 400% и более.
По характеру выполняемой производственной деятельности и величине энерготрат взрослое население может быть разделено на 5 групп (табл. 9.2).
Таблица 9.2
Суточный расход энергии в зависимости от характера выполняемой производственной деятельности
| Группа | Особенности профессии | Коэффициент физической активности | Суточный расход энергии, кДж (ккал) |
| Первая | Умственный труд | 1,4 | 9799 — 10265(2100 — 2450) |
| Вторая | Легкий физический труд | 1,6 | 10475 — 11732(2500 — 2800) |
| Третья | Физический труд средней тяжести | 1,9 | 12360 — 13827(2950 — 3300) |
| Четвертая | Тяжелый физический труд | 2,2 | 14246 — 16131(3400 — 3850) |
| Пятая | Особо тяжелый физический труд | 2,5 | 16131 — 17598(3850 — 4200) |
К первой группе относятся лица, занимающиеся умственным трудом, не требующим мышечных напряжений. Суточный расход у них составляет в среднем 2200—3000 ккал. У лиц, выполняющих механизированную работу, расход энергии повышен до 2350— 3200 ккал. При частично механизированном труде суточный расход энергии достигает 2500—3400 ккал. Очень тяжелый, немеханизированный физический труд вызывает расход энергии равный 2900— 3990 ккал. В отдельных случаях при выполнении длительной и тяжелой работы суточный расход достигает еще больших величин.
Спортивная деятельность сопровождается значительным увеличением суточного расхода энергии — до 4500—5000 ккал. В дни тренировок с повышенными нагрузками и в дни соревнований в некоторых видах спорта (лыжные гонки, бег на длинные дистанции и др.) эти величины могут быть еще больше.
Энерготраты при работе, которые рассчитывают на единицу времени или на единицу пути, прямо пропорциональны ее мощности. Суммарный же расход энергии зависит не только от мощности работы, но и от ее длительности. Например, при легкоатлетическом беге энерготраты, рассчитанные на 1 м пути, будут наибольшими при беге на 100 м, суммарный же расход энергии — наибольшим при марафонском беге.
На уровень энерготрат влияют температура и влажность воздуха, барометрическое давление, сила ветра, особенности грунта при беге, условия скольжения при беге на коньках и лыжах. Например, при одинаковой скорости бега на лыжах знерготраты при плохом скольжении будут значительно больше, чем при хорошем. На расход энергии влияют также эмоции, возникающие во время работы и, особенно при спортивной деятельности. Эмоции могут усиливать или, наоборот, снижать обмен веществ и энергии в организме.
При выполнении человеком механической работы коэффициент полезного действия может достигать 20—25%. Остальная освобождаемая в организме энергия превращается в тепло. Коэффициент полезного действия при мышечной деятельности зависит от структуры и темпа движений, от количества вовлекаемых в работу мышц и степени тренированности человека, выполняющего работу.
Оптимальный ритм движений, при котором энерготраты минимальны, различен при работе разной мощности. Он зависит также от тренированности человека.
Вопросы для самоконтроля