
Газообмен в тканях
В тканях кровь отдает кислород и поглощает углекислоту. Газообмен в
капиллярах тканей большого круга, так же как и в легочных капиллярах,
обусловлен диффузией вследствие разности парциальных напряжений газов в крови и
в тканях.
Напряжение углекислого газа в клетках может достигать 60 мм, в тканевой
жидкости оно весьма изменчиво и в среднем составляет 46 мм, а в притекающей к
тканям артериальной крови — 40 мм рт. ст. Диффунтируя по направлению более
низкого напряжения, углекислый газ переходит из клеток в тканевую жидкость и
далее в кровь, делая ее венозной. Напряжение углекислого газа в крови при
прохождении ее по капиллярам становится равным напряжению углекислого газа в
тканевой жидкости.
Клетки весьма энергично потребляют кислород, поэтому его парциальное
напряжение в протоплазме клеток очень низко, а при усилении их активности может
быть равно нулю. В тканевой жидкости напряжение кислорода колеблется между 20 и
40 мм. Вследствие этого кислород непрерывно поступает из артериальной крови,
приносимой к капиллярам большого круга кровообращения (здесь напряжение
кислорода равно 100 мм рт. ст.), в тканевую жидкость. В результате в оттекающей
от тканей венозной крови напряжение кислорода значительно ниже, чем в
артериальной, составляя 40 мм.
Кровь, проходя по капиллярам большого круга, отдает не весь свой кислород.
Артериальная кровь содержит около 20 об.% кислорода, венозная же кровь —
примерно 12 об. % кислорода. Таким образом, из 20 об. % кислорода ткани получают
8 об. %, т. е. 40% всего кислорода, содержащегося в крови.
То количество кислорода в процентах от общего содержания его в артериальной
крови, которое получают ткани, носит название коэффициента утилизации кислорода.
Его вычисляют путем определения разности содержания кислорода в артериальной и
венозной крови. Эту разность делят на содержание кислорода в артериальной крови
и умножают на 100.
Коэффициент утилизации кислорода меняется в зависимости от ряда
физиологических условий. В покое организма он равен 30—40%. При тяжёлой же
мышечной работе содержание кислорода в оттекающей от мышц венозной крови
уменьшается до 8—10 об. % и, следовательно, утилизация кислорода повышается до
50—60%.
Более быстрый переход кислорода в ткани обеспечивается раскрытием
нефункционировавших капилляров в работающей ткани. Повышению
коэффициента утилизации способствует также усиленное образование кислот —
молочной и угольной, что понижает сродство гемоглобина к кислороду и
обеспечивает более быструю диффузию кислорода из крови. Наконец, увеличению
утилизации кислорода содействуют повышение температуры работающих мышц и
усиление ферментативных и энергетических процессов, протекающих в клетках. Таким
образом, доставка кислорода к тканям регулируется в соответствии с
интенсивностью окислительных процессов.
Метаболические пути утилизации кислорода и продукция атф в ткани печени при острой циркуляторной гипоксии
Статья
Рис. 1. Показатели числа АОК селезенки мышей при ожоге с использованием риботана
Рис. 2. Показатели числа РОК селезенки мышей при ожоге с использованием риботана
1 т і
X’ ’ ‘ ■,(
1 /У ґ ‘
—і—- V /,
-І ■’ — -Ї
ч і
* » ■*
-0/F2.S 0/Р0.25 О/ГО,0S
Сутки эксперимента
Рис. 3. Показатели числа А-РОК селезенки мышей при ожоге с использованием риботана
Обобщая представленные данные, можно сказать о том, что ожоговая травма вызывает выраженные нарушения в гуморальном и клеточном звеньях иммунитета, выражающиеся в стойкой иммуносупрессии. Наиболее эффективно в качестве иммунокорректора риботан проявляет себя в дозах 0,25 и 2,5 мл/кг, а дозировка 0,05 мл/кг является недостаточной.
Литература
1. Азолов В.В и др. Актуальные вопросы патогенеза, клиники и лечения ожоговой болезни.- Горький, 1990.- С. 4-15.
2. Беляков И.М. и др. // Иммунол.- 1992.- № 4.- С. 22-25.
3. Хаитов Р.М., Пинегин Б.В. // Иммунол.- 2000.- № 5.— С. 4—7.
4. Яковлев Г М.и др. Резистентность, стресс, регуляция.— Л., 1990.— 267 с.
5. SparkesB.G. // Vaccine.— 1993.— P. 504—510.
УДК 616-005.4:612.26:612.35
МЕТАБОЛИЧЕСКИЕ ПУТИ УТИЛИЗАЦИИ КИСЛОРОДА И ПРОДУКЦИЯ АТФ В ТКАНИ ПЕЧЕНИ ПРИ ОСТРОЙ ЦИРКУЛЯТОРНОЙ ГИПОКСИИ
Т.П. ГЕНИНГ, Н.Н. ИВАНСКАЯ*
Одной из причин циркуляторной гипоксии является острая кровопотеря. Изучение отдельных звеньев патогенеза последствий кровопотери необходимо для успешного лечения постгемор-рагических состояний. При острой кровопотере всегда присутствуют факторы гипоксии и стресс-реакции [13]. При нормальных условиях эффективность биологического окисления соответствует функциональной активности органов и тканей. При нарушении этого соответствия возникает состояние энергетического дефицита, ведущее к разнообразным функциональным и морфологическим изменениям, направленным на формирование повышенной резистентности организма к гипоксическому воздействию [3].
Цель работы — изучение, в зависимости от объема кровопотери и сроков с момента воздействия, узловых параметров метаболизма кислорода и энергетического обмена в печени при острой циркуляторной гипоксии.
Материал и методы. Исследования проведены на беспородных белых крысах-самцах массой 190-210 граммов, содержавшихся в стандартных условиях вивария. Острую циркуляторную гипоксию вызывали по Sapirstein [15] кровопотерей объемами 1,5% и 0,75% от массы тела животного. Сроки с момента кровопотери до декапитации составили 3, 7 и 24 часа. В каждой экспериментальной группе использовали по 12 особей.
В гомогенате печени определяли активность цитохромок-сидазы (ЦХО) по Vernon, 1969 [2], активность сукцинатдегидро-геназы (СДГ) по Slater, 1959 [2], активность лактатдегидрогеназы (ЛДГ) по Меньшикову В.М., 1987 [9], концентрацию АТФ по Алейниковой Т.Л., 1988 [12]. Цифровые данные обрабатывали методом вариационной статистики с проверкой достоверности результатов по критерию Стьюдента и уровня значимости р<0,05.
Результаты. Установлено, что содержание АТФ у интакт-ных животных составило 4,092±0,15 мг/г ткани печени. Спустя 3, 7 и 24 часа с момента кровопотери объемом 1,5% концентрация АТФ составила 2,772±0,108 мг/г, 3,652±0,184 мг/г и 4,276±0,196 мг/г соответственно. При кровопотере объемом 0,75% на тех же сроках концентрация АТФ составила 2,618±0,165 мг/г; 2,748±0,182 мг/г и 3,966±0,235 мг/г (рис.1).
-1,50%
0,75%
Рис. 1. Содержание АТФ в ткани печени интактных крыс и на разных сроках после кровопотери объемами 1,5% и 0,75% от массы тела
На рис. 1 видно, что содержание АТФ в ткани печени снижается спустя 3 часа после кровопотери и восстанавливается до исходного уровня к 24 часам. При этом через 3 часа после гипок-сического воздействия концентрация АТФ независимо от объема кровопотери снижается на довольно близкую величину (32,26% и 36,02% от нормы), спустя 7 часов при более массивной кровопо-
ГОУ ВПО Ульяновский государственный университет, Институт Медицины, экологии и физической культуры, кафедра физиологии и патофизиологии. Россия, Ульяновск, ул. Ливчака.д.2, тел. (8422) 32-70-71
Т.П. Генинг, Н.Н. Иванская
тере (объемом 1,5%) содержание АТФ значительно выше, чем при меньшей кровопотере (объемом 0,75%).
Уровень активности ЦХО у интактных особей составил 18,70±056 мг/г ткани. После кровопотери объемом 1,5% активность фермента составила: спустя 3 часа 8,33±37 мг/г, через 7 часов — 18,68±1,12мг/Т, через 24 часа — 18,57±1,06 мг/г После кро-вопотери объемом 0,75% на тех же сроках активность ЦХО определялась как 13,32^0,80 мг/г, 18,51±1,07 мг/г и 18,73±093 мг/г соответственно (рис.2).
после кровопотери активность фермента достоверно увеличивается на 73,49%. При кровопотере объемом 0,75% активность фермента изменяется волнообразно. На сроках 3 и 24 часа активность ЛДГ достоверно повышается на 31,01% и 66,69% соответственно. На сроке 7 часов имеется тенденция к спаду активности фермента на 10,39% по отношению к интактным животным.
Рис. 2. Уровень активности ЦХО в ткани печени интактных крыс и на разных сроках после кровопотери объемами 1,5% и 0,75% от массы тела
В ходе исследования активности ЦХО выявлено достоверное снижение уровня активности фермента через 3 часа после кровопотери и возвращение к уровню интактных животных через 7 часов. Спустя 24 часа уровень активности ЦХО остается равным уровню интактных животных, независимо от объема крово-потери. Анализ уровня активности ЦХО через 3 часа после кро-вопотери показывает, что снижение активности фермента коррелирует с объемом кровопотери: при большем объеме потери крови степень снижения активности ЦХО меньше (55,46% при кровопотере 1,5 % и 28,74% при объеме 0,75%).
Установлено, что активность СДГ после кровопотери объемом 1,5% составила через 3 часа 395,0± 51,3 у/г, спустя 7 часов -325,0± 32,3 у/г, через 24 часа — 371,2± 37,3 у/г, что существенно превышает уровень активности фермента у интактных животных (250,0± 29,8 у/г ткани печени). Активность СДГ при кровопотере объемом 0,75% определялась как 639,2± 42,7 у/г, 203,9± 26,3 у/г и 411,6± 38,1 у/г через 3, 7 и 24 часа соответственно (рис.3).
700
B00
500
400
300
200
100
0
-1,50%
0,75%
Рис. З. Уровень активности СДГ в ткани печени интактных крыс и на разных сроках после кровопотери объемами 1,5% и 0,75% от массы тела
На рис. 3 видно, что уровень активности СДГ после крово-потери объемом 1,5% достоверно возрастает спустя 3 и 24 часа на 58,0% и 48,48% соответственно; через 7 часов имеется тенденция к увеличению активности фермента на 30,0% по сравнению с уровнем фермента у интактных животных. В группе животных с кровопотерей 0,75% от массы тела животного через 3 часа после воздействия уровень активности фермента достоверно возрастает через 3 и 24 часа на 155,66% и 64,59% соответственно; спустя 7 часов имеется тенденция к снижению на 18,44% по сравнению с уровнем активности фермента у интактных животных.
При исследовании активности ЛДГ уровень у интактных животных составил 122,2±1035 усл.ед./г ткани. При кровопотере объемом 1,5% активность фермента через 3, 7 и 24 часа после воздействия составила 212,Ш323 усл.ед./г, 129,&854
усл.ед./г,132,2±1206 усл.ед./г соответственно. После кровопотери объемом 0,75% соответственно указанным срокам активность ЛДГ определялась как 160,1±13,63 усл.ед./г, 109,5±10,88 усл.ед./г, 203,7^4,89 усл. ед./г (рис. 4).
По данным, уровень активности ЛДГ при кровопотере объемом 1,5% растет по отношению к интактным животным. На сроках 7 и 24 часа уровень активности ЛДГ имеет тенденцию к увеличению на 6,22% и 8,18% соответственно. Спустя 3 часа
Рис. 4. Уровень активности ЛДГ в ткани печени интактных крыс и на разных сроках после кровопотери объемами 1,5% и 0,75% от массы тела
Полученные биохимические показатели в печени крыс после воздействия острой кровопотери разного объема свидетельствуют о происходящих изменениях метаболических процессов в ткани. Исследуемые ферменты принимают непосредственное участие в клеточном метаболизме кислорода. Нарушение обеспечения клетки кислородом является следствием изменений системного кровообращения, сопровождающих острую потерю крови. Как известно, кислород — конечный акцептор электронов в дыхательной цепи, поэтому недостаток его приводит к уменьшению транспорта электронов и протонов, а следовательно, к рассогласованию субстратного окисления и фосфорилирования. Это приводит к уменьшению энергетического потенциала в ткани, выражающегося снижением концентрации АТФ, то есть эффективность аэробного энергетического обмена понижается [11].
Снижение содержания АТФ в условиях гипоксии показано М. В. Биленко [1]. Восстановление концентрации АТФ спустя 24 часа после кровопотери можно объяснить включением адаптационных механизмов по поддержанию процессов жизнеобеспечения. В связи с тем, что в настоящее время установлено, что АТФ разного происхождения (аэробного или анаэробного) потребляется в различных внутриклеточных процессах, вопрос о том, какие пути синтеза АТФ повреждены, а какие компенсаторно временно усилены или могут быть усилены медикаментозно, приобретает особое значение [1].
Компенсаторную функцию по синтезу дополнительных количеств АТФ в условиях гипоксии берут на себя анаэробные энергопродуцирующие процессы, важнейшим из которых является гликолиз [11]. Усиление гликолиза является частным проявлением компенсаторной перестройки метаболизма, направленным на поддержание уровня энергетического обеспечения клеток в условиях усиления в них АТФ-зависимых процессов. Стимуляция анаэробного гликолиза в печени выступает в роли универсальной реакции метаболизма на стресс [4]. В анаэробном гликолизе важную роль играет ЛДГ. Существует два пути реокисления внемитохондриального НАД, образующегося на окислительной стадии гликолиза: либо за счет пирувата с образованием лактата при участии внемитохондриальной ЛДГ, либо при участии дыхательной цепи митохондрий (челночный механизм). В анаэробных условиях лактат образуется в клетке вследствие того, что внеми-тохондриальный НАД-Н не может быть окислен дыхательной цепью и должен подвергнуться реокислению с участием пирува-та. В присутствии кислорода в большинстве клеток реокисление внемитохондриального НАД-Н должно происходить через дыхательную цепь, так как в обычных условиях лактат не накапливается в клетке. В аэробных условиях лактат может образовываться путем реокисления НАД-Н пируватом, если не действуют челночные системы, обеспечивающие поступление внемитохондри-ального НАД-Н в митохондрии. Накопление лактата способствует активации ЛДГ [7].
В литературе показано увеличение активности ЛДГ в процессе адаптации к гипоксическому воздействию согласуются с литературными [7]. Однако при летальном исходе циркуляторной гипоксии на криостатных срезах гистоэнзиматических реакций
Краткое сообщение
печени людей установлено снижение активности ЛДГ на 15,8% по сравнению с контролем [10]. Фосфорилирование с образованием АТФ происходит в нескольких участках дыхательной цепи, причем одним из важнейших является транспорт электронов через систему цитохромов [5]. Конечным компонентом цепи дыхательных ферментов, переносящим электроны от цитохрома на молекулярный кислород, является ЦХО. ЦХО катализирует более 95% утилизации клеточного кислорода. Уровень активности ЦХО может служить критерием выраженности гипоксии [8]. Согласно литературным данным [5] в печени крыс, погибавших при явлениях быстро наступившей декомпенсации кровообращения после кровопотери, активность ЦХО падала до 73% по сравнению с контрольным уровнем. Однако на ранних сроках после кровопотери (5 минут) активность ЦХО возрастала.
Результаты нашей работы по изучению уровня активности ЦХО частично согласуются с литературными. Выявленное снижение активности фермента через 3 часа после кровопотери и восстановление его активности через 7 часов свидетельствуют о произошедшей адаптации к возникшим условиям. При этом необходимо учитывать, что сильное снижение активности ЦХО наблюдается на терминальной стадии биоэнергетической гипоксии [8], развивающейся в условиях тяжелого и длительного экстремального воздействия с возможным летальным исходом. Одним из вариантов подобного состояния является полное прекращение кровотока в органе. В случае нашего эксперимента доза воздействия на орган не привела к летальному исходу, однако изменения уровня активности ЦХО были близки при разных объемах потери крови.
С дыхательной цепью митохондрий прочно связана СДГ. В опыте добавление СДГ к цитохромным частицам вызывает интенсивное дыхание, что объясняют восстановлением нативной структуры полиферментного комплекса [6]. В [1] также указывается, что при определенных условиях при ишемии в митохондриях наблюдается переход дыхания с НАД-зависимого на СДГ-зависимый путь. Кроме того, СДГ в значительной мере определяет скорость потребления кислорода и образования АТФ в дыхательной цепи. Уровень активности СДГ также может служить критерием оценки выраженности гипоксии [8].
По данным А.П. Довганского (1989) [5], в пределах 4 часов после кровопотери в печени крыс обнаружено возрастание активности СДГ. Рост активности СДГ в эксперименте А.П. Дов-ганский объясняет интенсивностью процессов в печени млекопитающих, значительной ролью в скорости потребления кислорода в дыхательной цепи и в адаптации организма к экстремальным воздействиям. В литературе есть опытные данные по двукратному увеличению активности СДГ и ЦХО в печени при кровопоте-ре; усилению окислительных процессов в печени в ближайшие сроки после геморрагии на фоне роста потребления кислорода; нормальной утилизации субстратов митохондриями печени при снижении сопряженности дыхания и фосфорилирования. (Mela, 1971) [14]. Оценка исследования корреляционной зависимости между уровнем ЦХО и СДГ, а также связи активности этих ферментов с содержанием АТФ в органе при разных условиях эксперимента (при n = 12), не обнаружили достоверных взаимосвязей. Результаты исследований свидетельствуют, что наибольшая степень выраженности изменений уровня активности ЦХО, СДГ, ЛДГ и концентрации АТФ в печени наблюдается через 3 часа с момента кровопотери, независимо от ее объема.
При этом снижение содержания АТФ в ткани, а затем восстановление через 24 часа после кровопотери можно объяснить изменениями путей синтеза макроэргических соединений при гипоксии. Так, на раннем сроке эксперимента наблюдается снижение активности ЦХО на фоне возрастания активности СДГ и ЛДГ. К 24 часам после кровопотери происходит адаптация к воздействию и восстановление работы дыхательной цепи, сопровождающееся нормализацией содержания АТФ и сохраняющегося небольшого повышения активности ЛДГ и СДГ по сравнению с контролем. Острая кровопотеря приводит к стадийному изменению параметров метаболизма кислорода разной степени выраженности, в зависимости от степени воздействия.
Литература
1. Биленко М.В. Ишемические и реперфузионные повреждения органов.- М., 1989.
2. Генинг Т.П. Ферментный спектр сыворотки крови и ткани печени при остром и хроническом токсическом поражении и в условиях стимуляции ее регенерации: Дис.. .канд.биол.наук.-Семипалатинск, 1980.
3. Гипоксические, гипокапнические и гиперкапнические состояния / Агаджанян Н.А. и др.- М., 2003.
4. Давыдов В.В, и др. // Патол. физиол. и эксперим. терапия.- 2005.- № 1.- С.12-14.
5. Довганский А.П. и др. Печень при экстремальных состояниях.- Кишинев, 1989.
6. Зимакова Н.И, Виноградов А.Д. // Митохондрии. Структура и функции в норме и патологии.- М., 1971.- С .189-195.
7. Ксейко Д.А. Антиоксидантная система печени и эритроцитов при вторичной тканевой гипоксии и в условиях коррекции аскорбиновой кислотой: Дис..канд. биол. наук.- Ульяновск, 2004.
8. Лукьянова Л.Д.// Вестн. РАМН.- 2000.- № 9.- С.9-12.
9. Меньшиков ВМ. Лабораторные методы исследования в клинике.- М., 1987.
10. Мишнев О.Д. и др. // Бюл. эксперим. биол. и мед.-1992.- Т. 113, № 4.- С. 435-437.
11. Резистентность к гипоксии/ Зиновьев Ю.В. и др.-Красноярск, 1988.
12. Руководство к практическим занятиям по биологической химии/ Алейникова Т.Л. и др.- М., 1988.
13. Утешев Н.С.и др.// Вестн. хирургии им. Грекова.-1999.- № 6.- С. 24-27.
14. Mela L., Bacalzo L.V., Miller L.D. // Amer. J. Phisiol.-1971.- Vol. 220.- P. 571-577.
15. Sapirstein R.A. at al.// Сй-c.Res.- 1960.- Vol. 8.- P. 135147.
THE OXIGEN METHABOLISM AND ATP CONCENTRATION IN A LIVER TISSUE AT ACUTE CIRCULATORY HYPOXIA
T.P. GENING, N.N. IVANSKAYA Summary
The dynamics of parameters of oxygen and energy metabolism in a liver tissue of rats is studied on the model of acute circulatory hypoxia at blood loss in volumes of 0,75 % and 1,5 % from the weight of a body. The activity of cytochromoxidase, succinate dehydrogenase, lactate dehydrogenase and the concentration of ATP are estimated in dependence on the terms from the moment of experimental influence and a degree of blood loss intensity. It is shown, that the most intensive changes are observed in 3 hours from the moment of the blood loss irrespective of its volume.
Key words: cytochromoxidase, succinate
Й Генинг Татьяна Петровна, зав.кафедрой физиологии и патофизиологии Института медицины, экологии и физической культуры Ульяновского госуниверситета, доктор биологических наук, профессор, академик РАЕН
ЙИванская Наталья Николаевна, аспирант кафедры физиологии и патофизиологии Института медицины, экологии и физкультуры Ульяновского госуниверситета, ассистент кафедры химии Ульяновского государственного политехнического университета
УДК 661.847.9
ОСОБЕННОСТИ ОБМЕНА ЦИНКА ПРИ ИНФАРКТЕ МИОКАРДА (КЛИНИКО-ЭКСПЕРИМЕНТАЛЬНОЕ ИССЛЕДОВАНИЕ)
А.Р. АНТОНОВ, Е.А. ВАСЬКИНА, Ю.Д. ЧЕРНЯКИН*
Цель — анализ уровня 7п в плазме крови больных инфарктом миокарда (ИМ) и в плазме крови и лимфе опытных животных линий Вистар и НИСАГ с экспериментальным ИМ (ЭИМ).
Методы исследования: обследовано 30 больных мужчин в острой стадии ИМ в сочетании с АГ, находящихся на лечении в отделении интенсивной терапии, контролем для которых служи-
* Новосибирский госмедуниверситет, г. Новосибирск
Пути утилизации кислорода клеткой
Большая часть кислорода, потребляемого клеткой (около 80 %), используется, как указано, в митохондриях с участием цитохромоксидазы. Это так называемый оксидазныйпуть. При этом происходит полное восстановление кислорода, причем субстрат не реагирует с кислородом непосредственно. Данный путь дает клетке энергию в виде АТФ. Помимо цитохромоксидазы существуют другие оксидазы (ФМН и ФАД-зависимые), которые катализируют реакции окисления веществ с образованием перекиси водорода
Наряду с этим существует другой путь окисления – оксигеназный. Он не дает клетке энергии, кислород включается в субстрат с образованием новой гидроксильной или карбоксильной группы. Этот путь происходит в основном в мембранах эндоплазматического ретикулума (микросомах). Путем микросомного окисления осуществляется α- и ω-окисление жирных кислот, синтез ненасыщенных жирных кислот, стероидов. Таким путем обезвреживаются ксенобиотики, т. е. чужеродные для организма вещества (лекарства, ядохимикаты, косметические препараты). Ферменты, осуществляющие такое окисление, называются оксигеназами. Различаютдиоксигеназы, которые включают в молекулу субстрата два атома молекулы кислорода. Более распространены в клетках монооксигеназы(гидроксилазы). Они катализируют реакции, при которых в молекулу субстрата включается один атом из молекулы кислорода, второй же атом кислорода восстанавливается при этом до воды. Монооксигеназные системы представляют собой короткие цепи переноса электронов и протонов, источником которых служит чаще всего восстановленный НАДФ , реже НАД или аскорбиновая кислота. Активатором кислорода при этом является цитохром Р450 – одноцепочечный хромопротеин с молекулярной массой 50 кДа.
Примерная схема монооксигеназной цепи выглядит следующим образом:
Смысл такого процесса заключается в том, что ксенобиотики, которые обычно гидрофобны, гидроксилируясь, становятся более гидрофильными, что способствует их обезвреживанию и выведению из организма с желчью или мочой. С участием микросомных систем осуществляется также биосинтез стероидов, желчных кислот, витамина Д3.
С появлением в атмосфере кислорода, а он появился тогда, когда возникли фотосинтезирующие организмы, стало возможным более эффективно использовать энергию, т. е. возник механизм окислительного фосфорилирования. Но, с другой стороны, вместе с этим кислород принес и новую опасность. При неполном восстановлении молекулы кислорода образуются высокоактивные формы (свободные радикалы), которые могут повреждать белки, нуклеиновые кислоты, липиды и способны даже убить живую клетку. Активные формы кислорода или свободные радикалы образуются в качестве промежуточных продуктов в ходе микросомного окисления, при работе дыхательной цепи, при воздействии ионизирующего излучения, при самопроизвольном окислении ряда веществ (гемоглобин). Свободные радикалы — молекулы, содержащие неспаренные электроны, агрессивные молекулы, которые атакуют другие молекулы с целью отнять у них электрон. К ним относятся: супероксидный радикал (О2–), перекисный радикал (О22–), гидроперекисный радикал (НО2–), пероксид водорода (Н2О2), гидроксильный радикал (НО–).
Свободные радикалы стимулируют разрывы в молекулах нуклеиновых кислот, нарушают функции белков, ведут к деполимеризации протеогликанов соединительной ткани, повреждают ненасыщенные жирные кислоты клеточных мембран, запуская, тем самым, механизм перекисного окисления липидов (ПОЛ). Вместе с тем свободные радикалы кислорода играют и положительную роль, так как участвуют в осуществлении фагоцитами и
Т-лимфоцитами их защитной функции.
Повышенное образование в организме свободных радикалов кислорода ведет к «окислительному стрессу», который может привести к повреждению мембран и гибели клетки. Поэтому в организме существуетантиоксидантная защита от свободных радикалов.
Различают неферментативную и ферментативную защиту клеток. Важнейшим компонентом неферментативной защиты является витамин Е (токоферол), витамин размножения.
Являясь жирорастворимым витамином, он всасывается вместе с липидами, поступает в лимфатическую систему и кровяное русло, а оттуда — в ткани. Токоферол защищает ненасыщенные жирные кислоты клеточных мембран от перекисного окисления, предохраняет от окисления SH-группы мембранных белков, защищает от окисления двойные связи в молекулах каротинов и витамина А. Токоферол (совместно с витамином С) способствует включению селена в состав активного центра глутатионпероксидазы — важнейшего фермента антиоксидантной защиты клеток. Он контролирует синтез гема, цитохромов, стабилизирует биологические мембраны.
Ферментативная защита клеток от свободных радикалов (антиоксидантная защита) осуществляется с помощью следующих ферментов:
· супероксиддисмутазы (превращает супероксидные радикалы в менее токсичную перекись водорода);
· каталазы (разлагает перекись водорода на воду и кислород);
· глутатионпероксидазы — главная система защиты эритроцитов от разрушительного действия перекиси водорода. В качестве кофермента глутатионпероксидаза использует трипептид – глутатион.
§
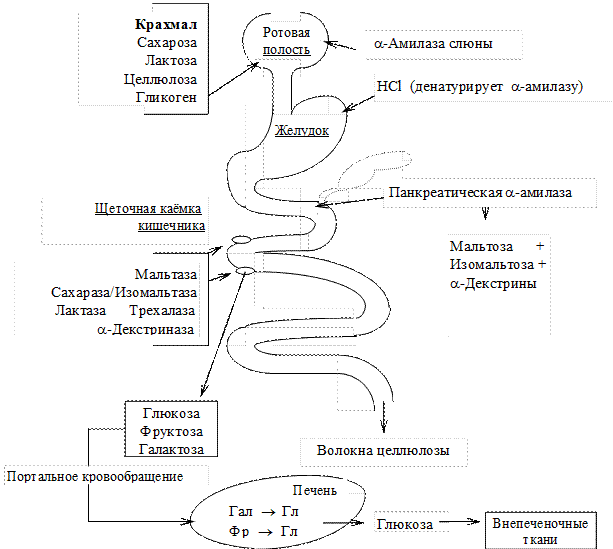
Ротовая полость. a-Амилаза слюны гидролизует внутренние a-1,4-гликозидные связи. Продуктами пищеварения являются олигосахаридные фрагменты (декстрины), в небольшом количестве — мальтоза и глюкоза.
Тонкий кишечник. Секретин стимулирует выделениепанкреатического сока.
Холецистокинин-панкреозимин стимулирует секрецию панкреатической a-амилазы и других панкреатических ферментов пищеварения.
Панкреатическая a-амилаза гидролизует внутренние a-1,4-гликозидные связи олигосахаридов и полисахаридов до мальтозы, изомальтозы и a-декстринов.
В ходе пристеночного пищеварения дисахаридазы гидролизуют дисахариды (мальтозу, изомальтозу, сахарозу, лактозу, трехалозу) до моносахаридов.
Мальтаза гидролизует мальтозу на две молекулы D-глюкозы.
Лактаза гидролизует лактозу на D-галактозу и D-глюкозу.
Изомальтаза/Сахараза — фермент двойного действия. Имеет два активных центра, расположенных в разных доменах. Фермент гидролизует сахарозу до D-фруктозы и
D-глюкозы, с помощью другого активного центра фермент катализирует гидролиз изомальтозы до двух молекул D-глюкозы.
Трехалаза гидролизует трехалозу на две молекулы D-глюкозы.
a—Декстриназа (терминальная декстриназа) также образуется в клетках слизистой кишечника и локализована в апикальной мембране энтероцитов. Фермент гидролизует
a-1,6-гликозидные связи в a-декстринах.
Роль системы утилизации кислорода тканями организма в аэробной производительности
Система тканевой утилизации кислорода включает в себя скелетные мышцы, сердце и дыхательные мышцы. К основным тканевым механизмам, совершенствующимся в процессе адаптации к различным факторам среды и увеличивающим способность ткани утилизировать кислород из крови, можно отнести следующие: увеличение числа и структуры митохондрий; повышение активности окислительных ферментов, в частности цитохро-
моксидазы, и др.; увеличение площади диффузионной поверхности в работающих мышцах за счет общего объема капилляров; вовлечение в деятельное состояние большего количества нейромоторных единиц; увеличение количества энергетических субстратов и миоглобина.
Индикация удельного вклада системы тканевой утилизации кислорода осуществляется по величине артериально-венозной разности по кислороду (АВР СЬ), что также ясно из уравнения Фика. Эта величина при предельной мышечной работе может достигать у нетренированных 120-140 мл на 1 л крови. У высококвалифицированных спортсменов этот показатель возрастает до 160-170 и даже 180 мл. Вполне понятно, что нарушение деятельности этой системы приводит к снижению производительности всей системы кислородного транспорта.
Таким образом, рассмотрев факторы, определяющие и лимитирующие аэробную производительность, следует заключить, что главным лимитирующим звеном в этой системе является сердечно-сосудистая система. Однако в качестве первичного звена, которое может создать затруднение в системе кислородного транспорта, может быть любая из ее составляющих (система вентиляции, система крови, система утилизации). В таком случае «удар» в конечном итоге будет нанесен сердечно-сосудистой системе, которая всегда служит конечно-лимитирующим звеном. Известным подтверждением этому является часто встречающаяся патология сердечнососудистой системы у спортсменов.