
: | Образовательная социальная сеть")
Взгляд снаружи
Диапазон концентрации кислорода в воздухе, пригодный для жизни. Диапазон содержания кислорода в воздухе ( p_{text{O}_2}), при котором возможна жизнедеятельность человека в течение длительного времени, ограничен значениями
90–100 мм рт. ст. < ( p_{text{O}_2}) < 400–450 мм рт. ст.
Нижняя граница соответствует началу кислородного голодания, верхняя — началу кислородного отравления. В процентном отношении наступление кислородного голодания у здорового человека наступает уже при содержании O2 в воздухе ( p_{text{O}_2}) / pатм менее 14% (при pатм = 760 мм рт. ст.).
Эти данные соответствуют диапазону жизнедеятельности человека на уровне моря. По мере подъема в горы давление снижается, что наглядно отражают кривые атмосферного давления и парциального давления кислорода (рис. 1).
Видно, что начиная с высот 4,5–5 км давление кислорода становится ниже допустимой нижней границы давления в 90 мм рт. ст. При этом давление воздуха в альвеолах составляет 105–110 мм рт. ст., что также близко к нижней границе. По мере уменьшения давления кислорода до уровня 100 мм рт. ст. замедляются обменные процессы в организме, дыхание и сердцебиение учащаются, ухудшаются зрение и работа мозга…
Оценка времени развития кислородной недостаточности при нахождении в замкнутом объеме. В качестве примера рассмотрим несколько ситуаций с людьми, находящимися в замкнутом объеме: один человек, застрявший в лифте объемом V = 2 м3; два человека в комнате с V = 30 м3; сто человек, застрявшие в остановившемся вагоне метро с V = 250 м3.
В каждом случае найдем, за какое время Δt в замкнутом объеме V в процессе спокойного дыхания людей концентрация кислорода снижается от первоначального уровня 21% до начала кислородной недостаточности, т.е. до 14%. Подчеркнем — спокойного, поскольку при панике это время сильно снижается.
Спокойному дыханию соответствует потребление кислорода на уровне 0,25 литра в минуту. Поскольку 1 литр O2 соответствует 5 ккал энергии, то 0,25 л/мин сообщает организму за сутки 0,25 × 5 × 60 × 24 ккал = 1800 ккал энергии. Так как плотность человеческого организма около 1000 кг/м3, тело массой 70 кг занимает объем 0,07 м3, или 70 литров. Добавив одежду, получим оценку объема, вытесняемого из замкнутого помещения, в 100 литров, или 0,1 кубометра на человека.
Лифт. Свободный объем, занятый воздухом, составляет 1,9 м3. В этом объеме содержится 1,9 × 0,21 м3 = 0,4 м3 = 400 л кислорода. Признаки кислородной недостаточности развиваются, когда полезный объем кислорода уменьшится до 1,9 × 0,14 м3 = 0,27 м3 = 270 л.
Комната. Свободный объем около 30 м3. Начальный объем кислорода 6,3 м3. Минимально допустимый объем кислорода 4,2 м3. Потребление кислорода 0,5 л/мин. Время ( Δt_{text{O}_2}) = 2100 / 0,5 мин = 4200 мин, т.е. почти трое суток (!).
Вагон метро. Свободный объем около 240 м3. Начальный объем кислорода 50 м3. Минимально допустимый объем кислорода 34 м3. Потребление кислорода около 25 л/мин . Время ( Δt_{text{O}_2}) = 16000/25 мин = 640 мин, т.е. около 10 часов.
Во всех указанных случаях (если нет паники) время развития кислородной недостаточности очень велико. Однако, такой вывод находится в противоречии с житейским опытом: в метро и застрявшем лифте бывает душно и даже после сна в комнате с закрытой форточкой наутро ощущается духота.
По всей видимости, имеет место другой, более мощный механизм развития неблагоприятных ощущений в процессе дыхания при нахождении в замкнутом объеме, не связанный с потерей кислорода из воздуха. Оказывается, таким механизмом является накопление углекислого газа.
Концентрация углекислого газа в воздухе, пригодная для жизни. Диапазон допустимого содержания CO2 в воздухе составляет
( 0 < C_{text{CO}_2} = frac{p_{text{CO}_2}}{p_{атм}} < text{0,1%}. )
Отметим, что обычное содержание углекислого газа в воздухе ( C_{text{CO}_2} ) = 0,04%.
Величину принятого ограничения сверху на содержание углекислого газа (( C_{text{CO}_{2:text{max}}} ) = 0,1%) обсудим чуть позже, а сначала проведем оценки для замкнутых объемов лифта, комнаты, вагона метро и школьного класса применительно ко времени накопления концентрации углекислого газа до верхней границы. Примем, что взрослый человек обычно выдыхает углекислого газа в атмосферу ( q_{text{CO}_2}) = 0,25 л/мин.
Лифт. Свободный объем, занятый воздухом, равен 1,9 м3. Изменение уровня содержания CO2 в воздухе от 0,04% до 0,1% займет
( Δt_{text{CO}_2} = frac{(C_{text{CO}_{2:text{max}}}:-:C_{text{CO}_2}):·:V}{q_{text{CO}_2}} = frac{(1:·:10^{-3}:-:4:·:10^{-4}):·:text{1,9}:·:10^3 }{text{0,25}}:text{мин} = 5:text{мин}. )
Комната. Свободный объем около 30 м3. Изменение уровня содержания CO2 в воздухе от 0,04% до 0,1% займет ( Δt_{text{CO}_2} ) = 6 · 10−4 · 30 · 103 / (2 · 0,25) мин = 36 мин.
Вагон метро. Свободный объем около 240 м3. Изменение уровня содержания CO2 в воздухе от 0,04% до 0,1% займет ( Δt_{text{CO}_2} ) = 6 · 10−4 · 240 · 103 / (100 · 0,3) мин ≈ 6 мин.
Школьный класс. Приведем также оценки для школьного класса объемом около 200 м3, в котором находится 25 учеников. При уровне выдоха CO2 одним школьником 0,12 л/м (половина от взрослого) получим ( Δt_{text{CO}_2} ) = 6 · 10−4 · 200 · 103 / (25 · 0,12) мин ≈ 40 мин.
Это уже ближе к житейским ощущениям и оправдывает присутствие вентиляции на потолке лифтов, необходимость проветривания комнат в домах, в школьных классах после каждого урока, а также наличие системы вентиляции в метро.
Таким образом, именно накопление углекислого газа в замкнутых помещениях в первую очередь действует угнетающе на человека. В чем это проявляется?
В литературе отмечается два типа воздействия: кратковременное (часы) и длительное (регулярно, более нескольких часов в день). Симптомы при кратковременном воздействии при уровне вдыхаемого углекислого газа выше 0,1% — это усталость, головная боль, ухудшение концентрации внимания, плохой сон…
При длительном воздействии при уровне CO2 выше 0,1% появляются проблемы с дыхательной системой (сухой кашель, риниты…), снижение иммунитета, ухудшение работы сердечно-сосудистой системы… При уровне выше 0,2% еще больше ухудшается концентрация внимания, растет количество совершаемых ошибок и т.д. по нарастающей.
Еще одна проблема помещений без вентиляции — возможность расслоения воздуха на фракции. Поскольку углекислый газ в полтора раза тяжелее воздуха, он может опуститься ближе к полу и его концентрация там увеличится. Но процесс этот медленный, и любое движение воздуха перемешивает фракции.
Наконец, использование растений, казалось бы, должно помочь — ведь они выделяют кислород и поглощают углекислый газ. Однако, это происходит только днем, а вечером и ночью (когда свежий воздух особенно нужен) растения выделяют углекислый газ, усугубляя проблему с его накоплением.
Накопление угарного газа в замкнутом помещении. Казалось бы, откуда взяться угарному газу (СО) в замкнутом помещении, если нет рядом дровяной печки или камина с неидеальной вытяжкой? Но в литературе приводятся следующие данные: наряду с углекислым газом человек выдыхает также и угарный газ — в количестве примерно 1,6 мл/ч (при нормальных условиях); предельно допустимая для человека концентрация угарного газа составляет 1 мг/м3.
Этих данных достаточно, чтобы снова провести оценки времени накопления предельной концентрации угарного газа для людей в лифте, комнате, вагоне метро и школьном классе. Для этого перейдем от объема к массе образовывающегося угарного газа, воспользовавшись известным соотношением: один моль любого газа при нормальных условиях занимает объем 22,4 л.
В таблице 2 приведены значения времени накопления CO2 и СО до опасной концентрации, а также времени развития кислородной недостаточности в лифте, комнате, вагоне метро и школьном классе. Для детей принята половинная величина выдыхаемого СО и CO2.
Таблица 2. Сопоставление времени снижения концентрации O2, накопления СО и CO2
Видно, что накопление углекислого газа примерно на порядок опаснее накопления угарного газа и еще на порядок опаснее снижения концентрации кислорода.
Мощность систем вентиляции. Как оценить мощность систем вентиляции qвент, необходимую для поддержания нормального состава воздуха? Если отвлечься от переходных процессов установления и выравнивания потоков воздуха, то конечный результат выглядит очень просто:
( q_{text{вент}} = frac{q_{text{CO}_2}}{(C_{text{CO}_{2:text{max}}}:-:C_{text{CO}_2})}. )
Так, если ( q_{text{CO}_2} ) = 0,25 литра в минуту (в этом случае человек выдыхает 15 литров CO2 в час), то при ( C_{text{CO}_{2:text{max}}} ) = 1 · 10−3 и ( C_{text{CO}_{2}} ) = 4 · 10−4 получим требуемую мощность вентиляции в 420 литров воздуха в минуту или 25 м3 в час.
Если же выдыхается 20 литров CO2 в час, то мощность вентиляции увеличивается до 33 м3 воздуха в час. А если принять для максимально допустимого значения концентрации CO2 в воздухе несколько меньшее значение 0,8 · 10−3, то мощность вырастет уже до 38 м3 воздуха в час (при 15 л CO2 в час) и 50 м3 воздуха в час (при 20 л CO2 в час).
Много это или мало? Как обеспечить такой приток свежего воздуха? Например, если приоткрыть дверь, то через каждый квадратный сантиметр щели при перепаде давлений по обе стороны двери Δp = 10 Па проходит в час один кубометр воздуха. Это означает, что при указанном Δp через сантиметровую щель в двери высотой два метра проходит 200 м3 воздуха за час.
Отметим, что принятый уровень перепада давлений 10 Па довольно мал (это 10−4 от атмосферного) и вполне может быть достигнут. Еще более мощный эффект вентиляции оказывает проветривание при открытии окон и дверей в течение хотя бы нескольких минут.
В качестве примера рассмотрим ситуацию с кислородом и углекислым газом при спасении детей в пещере Таиланда, частично затопленной водой. В 2022 году весь мир следил за спасением футбольной команды из 12 школьников и их тренера, ушедших на экскурсию в пещеру Кхао Луанг и застрявших в ней на 18 дней (23 июня — 10 июля) из-за дождей, затопивших вход в пещеру.
Они укрылись в воздушном кармане, полностью перекрытом водой и удаленном от выхода из пещеры на 5 километров. Задача заключалась в высвобождении ослабевших детей и тренера из пещеры. Ситуация осложнялась наличием узкой щели — на рисунке 2 она обозначена как «опасная точка», через которую предстояло выбираться.
В этой ситуации оказались важны все отмеченные выше особенности поведения кислорода и углекислого газа в замкнутом объеме. Для борьбы с постепенным уменьшением количества кислорода в пещере была организована доставка кислорода с помощью специального трубопровода.
Было решено, что накопление углекислого газа в пещере представляет существенно большую опасность, чем нехватка кислорода. Закачкой кислорода по трубопроводу в верхнюю часть пещеры вытесняли углекислый газ. Учитывалось также расслоение воздуха на фракции — CO2 скапливался в нижней части пещеры. Вот почему дети и тренер скрылись в верхней ее части.
Поиски ребят и подготовительные работы заняли почти две недели. За это время известный изобретатель и организатор исследований Илон Маск (космические корабли, электрокары) успел из запчастей к ракете изготовить миниатюрную подводную лодку на одного человека и доставить ее в Таиланд. Но из-за узкой щели от ее использования отказались.
Ситуация с каждым днем становилась все более сложной. Необходимо было постоянное присутствие людей, занятых на откачке воды из пещеры (иначе пещера полностью заполнилась бы водой) и установке труб для подачи кислорода. Более десятка аквалангистов доставляли в пещеру воду, еду и кислородные баллоны.
Там постоянно присутствовали врачи и те, кто готовили спасательную операцию. При дыхании этих взрослых спасателей состав воздуха ухудшался еще стремительнее. Наступил момент, когда из-за накопления углекислого газа дальше ждать было нельзя. Множество кислородных баллонов было расставлено по всему маршруту из пещеры к выходу (каждый баллон рассчитан на работу только в течение часа).
Тысяча спасателей снаружи, включая сто дайверов, начали операцию. В первый день 13 дайверов спасли четырех подростков. Во второй день 18 дайверов (и 70 аквалангистов сопровождения) спасли еще четверых. Наконец, в третий день были спасены оставшиеся четверо детей и их тренер, а также 4 человека, остававшиеся в пещере. Молодцы!
Некоторые современные сведения о факторах hif
Далее дорогой читатель будет вынужден простить мне несколько справочный раздел, суммирующий часть современных сведений о факторе HIF.
- Что есть гипоксия? Нормоксия — условие, при котором концентрация (напряжение) кислорода в окружающей организм/клетку среде «нормальное атмосферное», т.е. близко к 21%. Гипоксия, соответственно, это все, что ниже, гипероксия — все, что выше указанной цифры. На самом деле, понятия эти весьма условны. Классической нормоксии in vivo нет даже для дыхательного эпителия (~14,5%), а большинство тканей в норме прибывает в условиях так называемой физиологической гипоксии (2–9% в зависимости от ткани, обычно около 5%), которая для них и не гипоксия вовсе. Помимо этого, выделяют также умеренную гипоксию, собственно гипоксию и аноксию. Единой чёткой классификации гипоксий нет, но есть попытки (как в таблице 1).
- Как все начиналось. В самом начале протерозоя — 2,4 млрд. лет назад (или на 830 миллионов лет ранее, если верить совсем свежим данным [3]) — произошла так называемая кислородная катастрофа — довольно стремительное глобальное накопление кислорода в атмосфере Земли, что привело примерно 0,54 млрд. лет назад к формированию атмосферы, близкой к современной. Для многих организмов произошла смена восстановительного, анаэробного типа метаболизма на принципиально новый окислительный тип. Эволюционно возникла потребность в появлении кислород-чувствительного регулятора.
- Основные принципы работы кислород-чувствительных систем. Кислород-чувствительные системы прокариот и эукариот устроены сходным образом — в роли сенсора выступает фермент 2-оксоглутарат оксигеназа с двухвалентным железом в качестве кофактора [4]. Фермент в условиях нормоксии в присутствии 2-оксоглутара и молекулярного кислорода способен гидроксилировать остатки аминокислот (обычно пролина или аспарагина) в составе белка-регулятора транскрипции или трансляции, регулируя его функции (рис. 2).
Кислород-чувствительная система эукариот (PHD-HIF) претерпевала некоторую эволюцию. Уже довольно сходная с человеческой PHD-HIF система функционирует у Trichoplax adhaerens[5] — примитивного двухслойного животного, еще не обладающего билатеральной симметрией (рис. 3).
- Факторы HIFs и канонический кислород-зависимый путь регуляции их активности.У эукариот кислород-сенсорная система связана с функционированием важнейшего транскрипционного фактора HIF, регулирующего работу (по разным оценкам) 1–2% всех генов (что довольно приличная цифра). Факторы HIF представляет собой гетеродимерные белки, состоящие из конститутивно экспрессирующейся HIF-β субъединицы и одной из трех кислород-зависимых HIF-α субъединиц (HIF-1α, HIF-2α или HIF-3α), которые формируют при димеризации с β-субъединицами соответственно HIF-1, HIF-2 и HIF-3 (рис. 4). Лучше всего изучены факторы HIF-1 и HIF-2. Роль 2-оксоглутарат оксигеназы у эукариот выполняют пролилгидроксилазы (PHD 1-3), гидроксилирующие в условиях нормоксии остатки пролина в кислород-чувствительной α-субъединице, что приводит к ее деградации. В условиях гипоксии α- и β-субъединицы образуют гетеродимер, перемещаются в ядро и взаимодействуют с особыми последовательностями в промоторах генов HRE (hypoxia responsive elements), активируя экспрессию этих так называемых HIF-зависимых генов.
В условиях нормоксии транскрипционная активность HIF-1α и HIF-2α регулируется также за счет другого кислород-зависимого фермента — FIH-1 (factor inhibiting HIF-1), который, гидроксилируя остатки аспарагина, способен препятствовать взаимодействию HIFs с кофакторами [6]. - Неканонические пути регуляции активности факторов HIFs или та самая негипоксическая гипоксия. Помните пример с кобальтовой кардиомиопатией? Это был хороший пример так называемого неканонического пути активации HIF, когда он, по сути, активировался в условиях нормоксии. В данном случае Co2 выступает в качестве хелатора Fe2 (содержащегося в активном центре пролилгидроксилаз PHD), ингибирует их функцию и нарушает кислород-зависимое разрушение α-субъединицы. Это приводит к активации HIFs в условиях нормоксии. Так у любителей пива происходила постоянная активация HIFs и HIF-зависимого гена эритропоэтина — даже несмотря на то, что в их кровь была пересыщена гемоглобином, и никакой реальной гипоксии, разумеется, не наблюдалось. Воздействовать на Fe2 в составе пролилгидроксилаз PHD способны активные формы кислорода (АФК) [7], которые образуются в клетке в огромном количестве разнообразных процессов.
Позднее выяснилось, что активность HIF может регулироваться множеством других механизмов на уровне транскрипции, трансляции и посттрансляционных модификаций (фосфорилирование, ацетилирование/деацетилирование, убиквитинирование/деубиквитинирование, SUMO-илирование [8], S-нитрозилирование, NEDD-илирование). Механизмы эти могут быть как зависящими от кислорода, так и нет; как селективными, так и нет по отношению к HIF-1α и HIF-2α изоформам [9]. Но к этому мы вернемся несколько позже. - HIF-1 и HIF-2. В чем разница? А теперь еще один важный пункт — изоформоспецифические особенности фактора HIF. Напомню, что существуют главным образом две изоформы — HIF-1 и HIF-2; они обладают примерно 48%-идентичностью по аминокислотной последовательности, имеют один и тот же сайт для посадки — HRE — и, главным образом, один и тот же перечень контролируемых ими генов. Однако есть и особенности.
- Экспрессия первой изоформы повсеместна; экспрессия же HIF-2 тканеспецифична: это почечная ткань, кишечник, жировая ткань, эндотелий и другие.
- Существует специфика в активируемых изоформами генов: HIF-1 регулирует экспрессию ключевых ферментов гликолиза: 6-фосфофрукто-2-киназы/фруктозо-2,6-бифосфатазы 3, фосфоглицераткиназы, лактатдегидрогеназы А, карбангидразы-9; некоторых антиапоптотических факторов: BNIP3, BNIP3L; тогда как как HIF-2 (кстати, Семенза открыл именно фактор HIF-2) контролирует экспрессию «фактора стволовости» OCT-3/4 [10, 11], циклина D1, эритропоэтина и другие (рис. 5) [12].
- В настоящее время формируется парадигма кооперации двух факторов при гипоксии. Очень наглядно эта концепция продемонстрирована на модели динамики роста сосудов в ответ на ишемию и роста опухолевой массы (рис. 6). И в том, и в другом случае первоначально клетки находятся в состоянии выраженной гипоксии (аноксии) и активируются сразу оба фактора — HIF-1 и HIF-2, причем главным «игроком» является HIF-1. В это время происходят критические, быстрые события: стремительное переключение на гликолиз, защита от апоптоза, продукция ростовых факторов. Постепенно происходит переключение на изоформу HIF-2, которая активна при более умеренной гипоксии и более длительное время, экспрессируется другой спектр факторов, определяющих созревание сосудистой сети и дальнейший рост опухоли с метастазированием.
За переключение изоформ ответственна особая E3 лигаза, названная HAF — hypoxia-associated factor. В условиях продолжительной и незначительной гипоксии она способна связываться с С-концевым доменом HIF-2α, что приводит к его стабилизации и дополнительному повышению трансактивационных свойств HIF-2. В условиях острой, кратковременной гипоксии/аноксии активность ее подавлена. - Факторы HIF способны взаимодействовать с другими факторами, как связанными с регуляцией транскрипции (красивый пример — HIF-1 и HIF-2 по-разному взаимодействуют с участниками Notch-сигналинга и по-разному влияют на биологию стволовой клетки глиобластомы [13]), так и не связанными с транскрипцией (взаимодействие с g-секретазой при раке молочной железы [14]). Неканонические взаимодействия HIF-сигналинга, возможно, одна из самых интригующих и многообещающих областей.
- Случай из практики. В заключение данного раздела скажу, что явления негипоксической активации HIFs и различия в изоформной специфичности между HIF-1 и HIF-2 — это реальные вещи, с которыми я столкнулся лично. В своей дипломной работе я изучал экспрессию эндотелием различных хемокинов под действием ростовых факторов VEGF165 и HGF, по одиночке и в комбинации. Исследовалась закономерная активация киназ ERK-1/2, p38 и активация NF-kB. Экспрессия некоторых хемокинов и факторов адгезии (MCP-1, ICAM-1, VCAM-1) четко ложилась в нашу концепцию о «провоспалительном» действии VEGF165 и «противовоспалительном» действии HGF и согласовывалась с активностью провоспалительного фактора NFkB.
Однако экспрессия и продукция IL-8 (важнейшего провоспалительного хемокина) регулировалась загадочным и не совсем объяснимым образом. Позднее мы решили эту задачу и нашли, что в нашей системе фактор HGF способен стабилизировать и активировать HIF-2 в нормоксических условиях (скорее всего за счет прямого фосфорилирования HIF-2α и повышения стабильности) и, таким образом, повышать экспрессию и продукцию IL-8 (рис. 7), что и придавало загадочности в профиль его экспрессии [15].
Итак, дорогой читатель, я надеюсь, что вы не сильно утомились ознакомлением первой части рассказа. Скорее перейдем к его иммунологическому разделу!
Стабильность биосферы — чернова н.м., былова а.м. общая экология. учебник
Н.М. Чернова, А.М. Былова
Общая экология. Учебник
М.: Дрофа, 2004
Основой самоподдержания жизни на Земле являются биогеохимические круговороты. Процессы созидания органического вещества, аккумулирующего энергию, и противоположные процессы его разложения с высвобождением этой энергии одинаково необходимы для существования жизни. Все доступные для живых организмов химические соединения в биосфере имеют предел. Исчерпаемость пригодных для усвоения химических веществ часто тормозит развитие тех или иных групп организмов в локальных участках суши или океана. По выражению академика В. Р. Вильямса, единственный способ придать конечному свойства бесконечного – это заставить его вращаться по замкнутой кривой. Все химические элементы, используемые в процессах жизнедеятельности организмов, совершают постоянные перемещения, переходя из живых тел в соединения неживой природы и обратно. Возможность многократного использования одних и тех же атомов делает жизнь на Земле практически вечной при условии постоянного притока нужного количества энергии.
Глобальный биогеохимический круговорот вещества на планете представляет собой систему сложно переплетенных циклов отдельных элементов, связанных друг с другом по аналогии с деталями часового механизма (рис. 163). Объединяясь в разных пропорциях в составе сложных органических молекул, синтезируемых в живых телах, атомы различных элементов вне живого вещества имеют свою «судьбу» в атмосфере, гидросфере и литосфере. Круговороты планетарного масштаба создаются из бесчисленных локальных циклических перемещений атомов, движимых жизнедеятельностью организмов в отдельных экосистемах, и тех перемещений, которые вызываются действием ландшафтных и геологических причин (поверхностный и подземный сток, ветровая эрозия, движение морского дна, вулканизм, горообразование и т. п.). Различают поэтому малые и большие круговороты в соответствии с объемом рассматриваемых экосистем: локальные биологические круговороты, биогеохимические циклы материков и океанов и, наконец, общепланетарный биогеохимический круговорот.
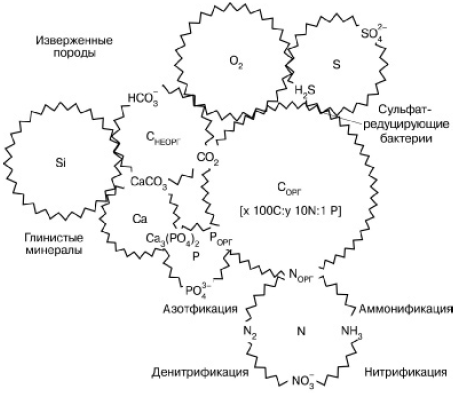
Рис. 163. Сопряжения циклов элементов в биосфере (по Г. А. Заварзину, 1998)
Вместе с тем отличительная черта биологических круговоротов – их неполная замкнутость. Часть химических элементов и их соединений постоянно выпадает из общей циркуляции и скапливается вне организмов, создавая своего рода запасы биогенных веществ. Так были накоплены кислород и азот в атмосфере, горючие ископаемые и другие породы земной коры. В масштабах геологического времени даже небольшое неравновесие в создании и разрушении органических веществ приводит к направленному преобразованию поверхностных слоев Земли.
Запасы биогенных веществ, мало доступные для живых организмов и медленно вовлекающиеся поэтому в биологический круговорот, составляют так называемый резервный фонд того или иного химического элемента в биосфере (например, кальция в карбонатных породах). Другая часть, интенсивно циркулирующая между телами организмов и окружающей средой, называется подвижным, или обменным, фондом. Соотношения между резервным и обменным фондами в биосфере у всех биогенных элементов различны.
Углерод. Углерод как химический элемент является основой жизни. Он может соединяться разными способами со многими другими элементами, образуя простые и сложные органические молекулы, входящие в состав живых клеток. По распространению на планете углерод занимает одиннадцатое место (0,35 % от веса земной коры), но в живом веществе в среднем составляет около 18 или 45 % от сухой биомассы.
В атмосфере углерод входит в состав углекислого газа СО2, в меньшей мере – в состав метана СН4 или следового количества других газообразных соединений. В гидросфере СО2 растворен в воде, и общее его содержание намного превышает атмосферное. Океан служит мощным буфером регуляции СО2 в атмосфере: при повышении в воздухе его концентрации увеличивается поглощение углекислого газа водой. Некоторая часть молекул СО2 реагирует с водой, образуя угольную кислоту, которая затем диссоциирует на ионы НСО3– и СО2-3. Эти ионы реагируют с катионами кальция или магния с выпадением карбонатов в осадок. Подобные реакции лежат в основе буферной системы океана, поддерживающей постоянство рН воды. При подкислении (увеличении концентрации ионов Н ) происходит сдвиг влево в цепи: СО2 воздуха → СО2 воды → Н2СО3 → Са(НСО3)2 → СаСО3. При подщелачивании усиливается выпадение в осадок карбонатов кальция.
Углекислый газ атмосферы и гидросферы представляет собой обменный фонд в круговороте углерода, откуда его черпают наземные растения и водоросли (рис. 164). Фотосинтез лежит в основе всех биологических круговоротов на Земле. Высвобождение фиксированного углерода происходит в ходе дыхательной активности самих фотосинтезирующих организмов и всех гетеротрофов – бактерий, грибов, животных, включающихся в цепи питания за счет живого или мертвого органического вещества.
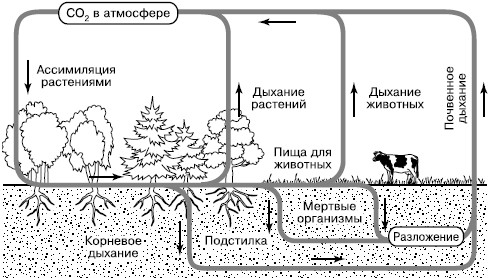
Рис. 164. Круговорот углерода в биосфере (по Б. Болину, 1972)
Особенно активно происходит возврат в атмосферу СО2 из почвы, где сосредоточена деятельность многочисленных групп деструкторов и редуцентов и осуществляется дыхание корневых систем растений. Этот интегральный процесс обозначается как «почвенное дыхание» и вносит существенный вклад в пополнение обменного фонда СО2 в воздухе. Параллельно с процессами минерализации органического вещества в почвах образуется гумус – богатый углеродом сложный и устойчивый молекулярный комплекс. Гумус является носителем почвенного плодородия, поскольку разрушается определенными группами микроорганизмов медленно и постепенно, обеспечивая равномерное питание растений. Гумус почв является одним из важных резервуаров углерода на суше.
В тех условиях, где деятельность деструкторов тормозится факторами внешней среды (например, при возникновении анаэробного режима в почвах и на дне водоемов), органическое вещество, накопленное растительностью, не разлагается, превращаясь со временем в такие породы, как каменный или бурый уголь, торф, сапропели, горючие сланцы и другие, богатые накопленной солнечной энергией. Они пополняют собой резервный фонд углерода, надолго выключаясь из биологического круговорота. Углерод временно депонируется также в живой биомассе, в мертвом опаде, в растворенном органическом веществе океана и т. п. Однако основным резервным фондом углерода на планете являются не живые организмы и не горючие ископаемые, а осадочные породы – известняки и доломиты. Их образование также связано с деятельностью живого вещества. Углерод этих карбонатов надолго захоранивается в недрах Земли и поступает в круговорот лишь в ходе эрозии при обнажении пород в тектонических циклах.
В биологическом круговороте участвуют лишь доли процента углерода от общего его количества на Земле. Углерод атмосферы и гидросферы многократно проходит через живые организмы. Растения суши способны исчерпать его запасы в воздухе за 4–5 лет, запасы в почвенном гумусе – за 300–400 лет. Основной возврат углерода в обменный фонд происходит за счет деятельности живых организмов, и лишь небольшая часть его (тысячные доли процента) компенсируется выделением из недр Земли в составе вулканических газов.
В настоящее время мощным фактором перевода углерода из резервного в обменный фонд биосферы становится добыча и сжигание огромных запасов горючих ископаемых.
По учетам в сети глобального мониторинга это уже приводит к повышению концентрации СО2 в атмосфере, последствия чего для судьбы человеческого общества усиленно обсуждаются и требуют научно обоснованного прогнозирования.
Кислород. С углеродным циклом теснейшим образом сопряжен круговорот кислорода.
Своей уникальной среди планет атмосферой с высоким содержанием свободного кислорода Земля обязана процессу фотосинтеза. Кислород освобождается из молекул воды и является по сути дела побочным продуктом фотосинтетической активности растений. Абиотическим путем кислород возникает в верхних слоях атмосферы за счет фотодиссоциации паров воды, но этот источник составляет лишь тысячные доли процента от поставляемыми фотосинтезом. Между содержанием кислорода в атмосфере и гидросфере существует подвижное равновесие. В воде его примерно в 21 раз меньше на равный объем по сравнению с воздухом.
Выделившийся кислород интенсивно расходуется на процессы дыхания всех аэробных организмов и на окисление разнообразных минеральных соединений. Эти процессы происходят в атмосфере, почве, воде, илах и горных породах. Показано, что значительная часть кислорода, связанного в осадочных породах, имеет фотосинтетическое происхождение. Обменный фонд О2 в атмосфере составляет не более 5 % от общей продукции фотосинтеза. Многие анаэробные бактерии также окисляют органические вещества в процессе анаэробного дыхания, используя для этого сульфаты или нитраты.
Накопление кислорода в атмосфере и гидросфере происходит в геологической истории в результате неполной замкнутости цикла углерода. На полное разложение органического вещества, создаваемого растениями, требуется точно такое же количество кислорода, которое выделилось при фотосинтезе. Захоронение органики в осадочных породах, углях, торфах послужило основой поддержания обменного фонда кислорода в атмосфере. Весь имеющийся в ней кислород проходит полный цикл через живые организмы примерно за 2000 лет.
В настоящее время значительная часть кислорода атмосферы связывается в результате работы транспорта, промышленности и других форм антропогенной деятельности. По расчетам, человечество тратит уже более 10 млрд т свободного кислорода из общего количества в 430–470 млрд т поставляемых процессами фотосинтеза. Если учесть, что в обменный фонд поступает лишь небольшая часть фотосинтетического кислорода, деятельность людей в этом отношении начинает приобретать угрожающие масштабы.
Вода. В ходе фотосинтеза растения используют водород воды в построении органических соединений, освобождая молекулярный кислород. В процессах дыхания всех живых существ, при окислении органических соединений вода образуется вновь. В истории жизни вся свободная вода гидросферы многократно прошла циклы разложения и новообразования в живом веществе планеты.
Кроме биологических циклов на Земле также осуществляется глобальный круговорот воды, движимый энергией Солнца. Вода испаряется с поверхности водоемов и суши и затем вновь поступает на Землю в виде осадков. Над океаном испарение превышает осадки, над сушей – наоборот. Эти различия компенсируются речным стоком. В глобальном круговороте воды растительность суши играет немаловажную роль. Транспирация растений на отдельных участках земной поверхности может составить до 80–90 % выпадающих здесь осадков, а в среднем по всем климатическим поясам – около 30 %. В истории биосферы появление наземной растительности вызвало за счет транспирации перераспределение осадков над морем и сушей и тем самым косвенно повлияло на размеры речного стока и процессы физического и химического выветривания.
Азот. В атмосфере и живом веществе содержится менее 2 % всего азота на Земле, но именно он поддерживает жизнь на планете. Азот входит в состав важнейших органических молекул – ДНК, белков, липопротеидов, АТФ, хлорофилла и других. В растительных тканях его соотношение с углеродом составляет в среднем 1: 30, а в морских водорослях 1: 6. Биологический цикл азота поэтому также тесно связан с углеродным.
Молекулярный азот атмосферы недоступен растениям, которые могут усваивать этот элемент только в виде ионов аммония, нитратов или из почвенных или водных растворов. Поэтому недостаток азота часто является фактором, лимитирующим первичную продукцию. Тем не менее атмосферный азот широко вовлекается в биологический круговорот благодаря деятельности прокариотических организмов (рис. 165). Способностью к фиксации молекулярного азота обладают очень многие прокариоты. В большой мере она развита у фотосинтезирующих сине-зеленых водорослей (цианобактерий). Активно фиксируют азот свободно живущие в почве бактерии рода Azotobacter, а также клубеньковые бактерии Rhizobium, живущие на корнях растений семейства бобовых. При этом может связываться до 400 кг азота на га в год. Отмирая, бактериальные клетки обогащают почву азотными соединениями, доступными для растений. Симбиотические формы снабжают хозяина и за счет прижизненных выделений.
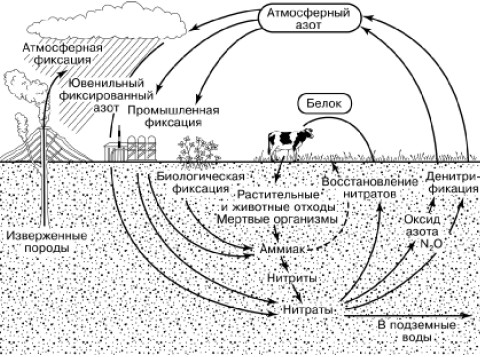
Рис. 165. Круговорот азота в биосфере (по К. Делвичу, 1972)
В круговороте азота принимают большое участие также аммонифицирующие микроорганизмы. Они разлагают белки и другие содержащие азот органические вещества до образования аммиака. В аммонийной форме азот частью вновь поглощается корнями растений, а частью перехватывается нитрифицирующими микроорганизмами. Они используют для себя химическую энергию, окисляя аммиак сначала в нитритную форму (бактерии Nitrosomonas). Окисление нитритов в нитраты производят бактерии рода Nitrobacter. Образовавшиеся нитраты вновь используются растениями в ходе фотосинтеза.
Прямо противоположна по функциям группа микроорганизмов – денитрификаторов.
В анаэробных условиях в почвах или водах они используют кислород нитратов для окисления органических веществ, получая энергию для своей жизнедеятельности. Азот при этом восстанавливается до молекулярного. Азотфиксация и денитрификация в природе приблизительно уравновешены. Цикл азота, таким образом, зависит преимущественно от деятельности бактерий, тогда как растения встраиваются в него, используя промежуточные продукты этого цикла и намного увеличивая масштабы азотной циркуляции в биосфере за счет продуцирования своей биомассы. Объемы микробной фиксации азота составляют до 2,5 т на км2 в год.
Небиологическая фиксация азота и поступление в почвы его окислов и аммиака происходит также с дождевыми осадками при ионизации атмосферы и грозовых разрядах. В среднем это дает около 1 т связанного азота на км2 в год.
Современная промышленность удобрений фиксирует азот атмосферы в размерах, превышающих природную азотфиксацию в целях увеличения продукции сельскохозяйственных растений.
Высокие дозы азотных удобрений приводят, однако, к вымыванию нитратов в грунтовые воды, водоемы и в конечном счете – питьевую воду, а также к избытку их в продуктах питания, что является опасным для человека. Таким же источником загрязнений служат сточные воды с высоким содержанием аммония. На его окисление до нитратов тратится растворенный в воде кислород, что часто бывает губительно для гидробионтов. Таким образом, деятельность человека все сильнее влияет на круговорот азота, в основном в сторону превышения перевода его в связанные формы над процессами возврата в молекулярное состояние.
Фосфор. Этот элемент, необходимый для синтеза многих органических веществ, включая АТФ, ДНК, РНК, усваивается растениями только в виде ионов ортофосфорной кислоты (РО3 4). Он относится к элементам, лимитирующим первичную продукцию и на суше, и особенно в океане, поскольку обменный фонд фосфора в почвах и водах невелик. Круговорот этого элемента в масштабах биосферы незамкнут.
На суше растения черпают из почвы фосфаты, освобожденные редуцентами из разлагающихся органических остатков, но в щелочной или кислой почве растворимость фосфорных соединений резко падает. Основной резервный фонд фосфатов содержится в горных породах, созданных на дне океана в геологическом прошлом. В ходе выщелачивания пород часть этих запасов переходит в почву и в виде взвесей и растворов вымывается в водоемы. В гидросфере фосфаты используются фитопланктоном, переходя по цепям питания в других гидробионтов. Однако в океане большая часть фосфорных соединений захоранивается с остатками животных и растений на больших глубинах и не попадает вновь в фотическую зону, переходя с осадочными породами в большой геологический круговорот. На глубине растворенные фосфаты связываются с кальцием, образуя фосфориты и аппатиты. В биосфере, по сути дела, происходит однонаправленный поток фосфора из горных пород суши в глубины океана, и обменный фонд его в гидросфере очень ограничен. Биологическая продуктивность океана резко повышается в районах, где увеличивается доступность фосфора для фитопланктона – при сносе его речными водами, у побережий морей и в зонах так называемого апвеллинга – восходящих океанических течений, возвращающих фосфорные соединения со дна в освещенные слои воды.
Наземные залежи фосфоритов и аппатитов разрабатываются в качестве удобрений. Снос избыточного фосфора в пресные водоемы является одной из главных причин их «цветения» – бурной эвтрофикации.
Сера. Круговорот серы, необходимой для построения ряда аминокислот, ответственных за трехмерную структуру белков, поддерживается в биосфере широким спектром бактерий. В отдельных звеньях этого цикла участвуют аэробные микроорганизмы, окисляющие серу органических остатков до сульфатов, а также анаэробные сульфатредукторы, восстанавливающие сульфаты до сероводорода, кроме них – разные группы серобактерий, окисляющих сероводород до элементарной серы и далее – сульфатов, тионовые бактерии, переводящие элементарную серу также в соли серной кислоты. Растения усваивают из почвы и воды только ионы SO2-4, поставляемые им деятельностью прокариотов.
Основное накопление серы происходит в океане, куда сульфатные ионы непрерывно поступают с суши с речным стоком. Частично сера возвращается в атмосферу при выделении из вод сероводорода и окисляется здесь до двуокиси, превращаясь в дождевой воде в серную кислоту. Промышленное использование большого количества сульфатов и элементарной серы и сжигание горючих ископаемых поставляют в атмосферу большие объемы диоксида серы. Это вредит растительности, животным, людям и служит источником кислотных дождей, усугубляющих отрицательные эффекты вмешательства человека в круговорот серы.
Как мы видим, круговороты всех биогенных элементов на планете поддерживаются сложным взаимодействием разных частей биосферы. Они формируются деятельностью разных по функциям групп организмов, системой стока и испарения, связывающих океан и сушу, процессами циркуляции вод и воздушных масс, действием сил гравитации, тектоникой плит и другими масштабными геологическими и геофизическими процессами. Биосфера действует как единая сложная система, в которой циркулируют с разной скоростью атомы отдельных элементов. Однако главным двигателем этих круговоротов является живое вещество планеты, все живые организмы, обеспечивающие процессы синтеза, трансформации и разложения органического вещества.
Вместо заключения
В настоящее время разработаны/разрабатываются методы селективного ингибирования HIF-1 и HIF-2; селективного ингибирования различных изоформ пролилгидроксилаз PHDs, за счет чего возможно активировать HIF-1 и HIF-2, причем также селективно; можно ингибировать фермент FIH-1и прочие участники пути регуляции кислород-зависимого пути деградации. При желании и достаточной фантазии можно пробовать воздействовать на кислород-независимые пути.
Все это, несомненно, должно найти и, я уверен, найдет применение в практической медицине. Но это потребует крайней обдуманности, многостадийного контроля и досконального изучения. Чем далеко ходить, лучше приведу пример.
Не так давно, в начале-середине 2000-х годов, наблюдался некоторый бум: для многих типов онкологических заболеваний была показана сверхэкспрессия HIF-1α, что вполне соответствовало понятиям об опухолевой биологии: быстрорастущая опухолевая масса в условиях жесткой гипоксии переходит на гликолитический анаэробный тип метаболизма [23], при этом активно секретируя вокруг себя многочисленные факторы роста сосудов, факторы инвазии и т.д. Что делать? Подавим экспрессию HIF-1α и дело в шляпе! Не тут-то было — реальность оказалась сложнее и запутанней.
Так, например, при применении siRNA против HIF-1α на культуре пигментного эпителия сетчатки и эндотелия сосудов, наблюдалось вполне закономерное снижение секреции таких ангиогенных факторов как VEGF, TGF-β (это очень хорошо), но росла секреция IL-8, мощного хемокина с ярко выраженными ангиогенными свойствами (это очень плохо)
[24]. Позднее, группой профессора Лобода был раскрыт механизм — дело в том, что HIF-1 подавляет экспрессию IL-8, а HIF-2 — активирует. Подавляя HIF-1 в клетках, исследователи добивались реципрокной активации HIF-2 и экспрессии IL-8 [25]. Такая неоднозначная выходила терапия.
Также нужно быть аккуратными с иммунной системой. Системное подавление HIF-1 при аутоиммунных заболеваниях, возможно, и приведет к снижению популяции Th17 и росту числа Treg, что теоретически способно облегчить течение заболевания, но также способно привести к искусственному комбинированному иммунодефициту за счет дисфункции M1-макрофагов, нейтрофилов, Th1, Th2, Th17 и СD8 T лимфоцитов.
Это, конечно, спекуляции, но все же.
- Alexander C.S. (1972). Cobalt-beer cardiomyopathy: a clinical and pathologic study of twenty-eight cases. Am. J. Med. 53 (4), 395–417;
- Gregg L. Semenza, M.D., Ph.D.Сайт The Johns Hopkins Hospital;
- Satkoski A.M., Beukes N.J., Li W., Beard B.L., Johnson C.M. (2022). A redox-stratified ocean 3.2 billion years ago. Earth Planet. Sci. Lett. 430, 43–53;
- Scotti J.S., Leung I.K., Ge W., Bentley M.A., Paps J., Kramer H.B. et al. (2022). Human oxygen sensing may have origins in prokaryotic elongation factor Tu prolyl-hydroxylation. Proc. Natl. Acad. Sci. USA.111 (37), 13331–13336;
- Loenarz C., Coleman M.L., Boleininger A., Schierwater B., Holland P. W., Ratcliffe P.J., Schofield C.J. (2022). The hypoxia‐inducible transcription factor pathway regulates oxygen sensing in the simplest animal, Trichoplax adhaerens. EMBO Rep. 12 (1), 63–70;
- Prabhakar N.R. and Semenza G.L. (2022). Oxygen sensing and homeostasis. Physiology. 30 (5), 340–348;
- Активный кислород: друг или враг, или О пользе и вреде антиоксидантов;
- SUMO: японская борьба или уникальная посттрансляционная модификация?;
- Agani F. and Jiang B.H. (2022). Oxygen-independent regulation of HIF-1: novel involvement of PI3K/AKT/mTOR pathway in cancer. Curr. Cancer. Drug Targets. 13 (3), 245–251;
- Была клетка простая, стала стволовая;
- Ствол и ветки: стволовые клетки;
- Koh M.Y. and Powis G. (2022). Passing the baton: the HIF switch. Trends Biochem. Sci. 37 (9), 364–372;
- Hu Y.Y., Fu L.A., Li S.Z., Chen Y., Li J.C., Han J. et al. (2022). Hif-1α and Hif-2α differentially regulate Notch signaling through competitive interaction with the intracellular domain of Notch receptors in glioma stem cells. Cancer Lett. 349 (1), 67–76;
- Villa J.C., Chiu D., Brandes A.H., Escorcia F.E., Villa C.H., Maguire W.F. et al. M. (2022). Nontranscriptional role of Hif-1α in activation of γ-secretase and notch signaling in breast cancer. Cell Rep. 8 (4), 1077–1092;
- Gluhanyuk E., Makarevich P., Gallinger J., Dergilev K., Beloglazova I., Parfyonova Ye. (2022). Diverse modulation of endothelial chemokine production by VEGF165 and HGF via NFkB and HIF-2. Материалы конференции Hypoxia: From Basic Mechanisms to Therapeutics;
- Palazon A., Goldrath A.W., Nizet V., Johnson R.S. (2022). HIF transcription factors, inflammation, and immunity. Immunity. 41 (4), 518–528;
- Phan A.T. and Goldrath A.W. (2022). Hypoxia-inducible factors regulate T cell metabolism and function. Mol. Immunol. doi: 10.1016/j.molimm.2022.08.004;
- Hsiao H.W., Hsu T.S., Liu W.H., Hsieh W.C., Chou T.F., Wu Y.J. et al. (2022). Deltex1 antagonizes HIF-1α and sustains the stability of regulatory T cells in vivo. Nat. Commun. 6, 6353;
- Yao Y., Vent-Schmidt J., McGeough M.D., Wong M., Hoffman H.M., Steiner T.S., Levings M.K. (2022). Tr1 cells, but not Foxp3 regulatory T cells, suppress NLRP3 inflammasome activation via an IL-10—dependent mechanism. J. Immunol. 195 (2), 488–497;
- Netea M.G., Latz E., Mills K.H., O’Neill L.A. (2022). Innate immune memory: a paradigm shift in understanding host defense. Nat. Immunol. 16 (7), 675–679;
- Одураченные макрофаги, или Несколько слов о том, как злокачественные опухоли обманывают иммунитет;
- Thomas A., Tambuwala M.M., McNicholas W.T., Roche H.M., Taylor C.T., Pepin J.L. et al. (2022). Chronic intermittent hypoxia contributes to pro-inflammatory macrophage alteration in visceral adipose tissue of lean and obese mice. Am. J. Respir. Crit. Care Med. 191, A2691;
- Страшней клешней на свете нет…;
- Forooghian F. and Das B. (2007). Anti-angiogenic effects of ribonucleic acid interference targeting vascular endothelial growth factor and hypoxia-inducible factor-1alpha. Am. J. Ophthalmol. 144 (5), 761–768;
- Loboda A., Jozkowicz A., Dulak J. (2022). HIF-1 versus HIF-2 — Is one more important than the other?Vascul. Pharmacol. 56 (5), 245–251..