

Роль кислорода в жизнедеятельности рыб – аграрий
Все рыбы дышат растворенным в воде кислородом, поэтому его содержание для них имеет решающее значение. Дышать водным животным значительно тяжелее и не только потому, что в воде в 21 раз меньше кислорода, но и потому, что вода плотнее воздуха в 800 раз.
Пелагические, речные и холодолюбивые рыбы более требовательны к кислороду, чем донные.
Кислород необходим рыбам для обеспечения аэробного энергообмена в индивидуальном развитии, и они могут обходиться без него самое короткое время только на ранних стадиях. Гликолиз у рыб чаще всего имеет место в зрелых половых клетках и у эмбрионов, т.е. в самом начале становления новых организмов.
В подавляющем большинстве рыбы используют кислород, растворенный в воде, и лишь некоторые виды способны дополнительно использовать атмосферный кислород .
По отношению к кислороду рыб делят на следующие группы:
- нуждающиеся в высоком содержании кислорода (7-12 мг/л), при снижении его содержания до 5-6 мг/л дыхание невозможно (форели, сиги);
- нуждающиеся в высоком содержании кислорода (5-8 мг/л), но выдерживающих его уменьшение до 5 мг/л (многочисленная группа пресноводных рыб: хариус, подуст, пескарь, налим);
- менее требовательные к содержанию кислорода, легко переносящие его уменьшение до 5 мг/л (окунь, карп, плотва, щука);
- довольствующиеся содержанием кислорода в 2,0-0,5 мг/л (линь, сазан, карась).
Морские рыбы более чувствительны к понижению содержания кислорода, чем речные, и задыхаются при уменьшении его содержания до 60-70% нормы (Котляр, 2007).
Потребление кислорода рыбами зависит от вида, возраста, подвижности, плотности посадки, физиологического состояния и солености воды.
Молодь рыб более чувствительна к содержанию кислорода, чем старшие возрастные группы.
Подвижные рыбы больше потребляют кислорода, чем малоподвижные.
Перед нерестом потребление кислорода рыбами возрастает на 23-30% по сравнению с другими периодами.
В холодной воде кислорода растворяется больше, чем в теплой, следовательно, при низких температурах рыба нуждается в меньшем количестве гемоглобина.
Рыбам вреден не только недостаток кислорода, но и его избыток который вызывает анемию и удушье.
Обогащение воды кислородом происходит в основном двумя путями: продуцированием кислорода фтосинтезирующими растениями и поступлением его из атмосферы.
Расходуется кислород на обеспечение процессов жизнедеятельности гидробионтов и окисление органических и минеральных веществ. Следовательно, любые воздействия на водоем, которые снижающие продуцирование кислорода или увеличивающие его расход, могут принести к нарушению кислородного режима водоема, к возникновению в нем кратковременного или длительного дефицита.
Даже в нормальных условиях концентрация растворенного кислорода в пресных водоемах претерпевает значительные изменения в зависимости от интенсивности фотосинтеза и степени насыщения воды воздухом. В теплых поверхностных слоях, где фотосинтез идет особенно интенсивно, концентрация кислорода, как правило, выше, чем в более глубоких слоях. В морях и океанах, где перемешивание более эффективно, концентрация кислорода и углекислого газа более постоянна, чем в пресных водоемах.
Существенное влияние на уровень насыщения воды кислородом оказывает температура, поскольку с ее изменениями меняется величина растворимости кислорода. При прочих равных условиях растворимость кислорода в пресной воде выше, чем в соленой.
Помимо температуры и солености, на содержание кислорода в воде влияют сезонные и суточные изменения интенсивности фотосинтеза водных растений, особенности динамики потребления кислорода рыбами и другими водными животными, количество находящихся в воде легко окисляемых органических и минеральных веществ, сезонные особенности поглощения кислорода поверхностными слоями воды из воздуха. Вследствие этого в водоеме имеют место сезонные и суточные колебания концентрации растворенного в воде кислорода.
Кратковременные суточные изменения концентраций кислорода в воде представляют для рыб меньшую опасность, чем более длительные сезонные изменения. Зимой, вследствие образования ледяного покрова, препятствующего поступлению кислорода из воздуха, содержание растворенного в воде кислорода во многих водоемах снижается до 50–25% нормы по сравнению с летним периодом .
Дефицит кислорода приводит к массовым заморам рыб, нередко заморы возникают летом, главным образом ночью, из-за усиленного потребления кислорода водной растительностью или при массовом отмирании водорослей, чаше всего в слабо проточных водоемах.
Еще более существенное воздействие на кислородный режим водоемов оказывают загрязнения, поступающие с промышленными, сельскохозяйственными и бытовыми сточными водами. Большинство сточных вод, наряду с прямым токсическим воздействием на рыбу, вызывает резкий дефицит растворенного в воде кислорода, ведущий к обеднению кормовой базы и исчезновению оксифильных видов рыб (Котляр, 2007).
Около половины всех случаев массовой гибели рыб в загрязненных водоемах обусловлено резким дефицитом растворенного в воде кислорода, в связи с усиленным его расходом на окисление органических загрязнений.
Устойчивость рыб к дефициту кислорода определяется глубиной и длительностью наступивших изменений, температурой воды, ее химическим составом, видовыми и возрастными особенностями рыб.
Интенсивность потребления рыбой кислорода находится в прямой зависимости от температуры воды. При низких температурах потребность рыб в кислороде меньше, чем при высоких.
Чувствительность рыб к недостатку кислорода у холодолюбивых рыб значительно выше, чем у теплолюбивых (карповых, окуневых), а устойчивость, напротив, у теплолюбивых выше, чем у холодолюбивых, хотя разрыв между пороговым напряжением кислорода, вызывающим гибель и критическим, при котором наступает реальное угнетение дыхания и снижение потребление кислорода, чрезвычайно мал, что делает рыб более уязвимыми при резком изменении кислородного режима.
Рыбы способны выживать при концентрации кислорода ниже уровня нормального насыщения. Длительность выживания определяется степенью снижения содержания кислорода: чем существеннее отклонения, тем короче время выживания и, наоборот, чем менее значительны изменения концентрации кислорода, тем длительнее время выживания в дискомфортных условиях.
Хотя рыбы и способны переносить низкие концентрации кислорода более или менее длительный период не погибая, почти любое снижение содержания кислорода ниже уровня насыщения отрицательно влияет на рост и воспроизводство и другие физиологические функции рыб. Особенно велико отрицательное влияние пониженных концентраций кислорода на ранних этапах развития и роста рыб.
При недостаточном содержании кислорода возникают различные нарушения в строении зародышей рыб. При снижении кислорода в воде, мальки не могут заполнить плавательный пузырь воздухом, подняться на плав и начать питаться. При этом вылупившиеся личинки имеют меньшую массу и размеры в сравнении с личинками, развитие которых проходило при нормальном насыщении кислорода. Дальнейшее снижение концентрации кислорода заканчивается гибелью всех зародышей еще до завершения инкубации.
Низкое содержание кислорода в воде (0,5-3,0 мг/л) оказывает губительное действие на большинство видов рыб. При содержании в воде растворенного кислорода ниже 4 мг/л многие промысловые виды рыб испытывают затруднения в дыхании, а у лососевых и осетровых угнетение дыхания может наступить даже при концентрации кислорода ниже 6 мг/л.
Большую угрозу для жизни рыб представляют сточные воды, содержащие бистро и медленно окисляющиеся вещества, которые весьма интенсивно поглощают кислород, вызывая тем самым снижение его содержания в водоемах. Особенно опасны в этом отношении хозяйственные сточные воды и стоки пищевых предприятий (мясокомбинатов, сахарных и картофелекрахмальных, винокуренных, дрожжевых) и кожевенных заводов. Обилие органики животного и растительного происхождения в сточных водах этих предприятий, как правило, лишенных специфических токсических свойств, ведет к отложению их на дне и формированию донных отложений.
Органические донные отложения со временем подвергаются процессам гниения, брожения и окисления. Эти процессы связаны с расходованием огромного количества кислорода, что приводит к резкому снижению его содержания в воде. Не меньшую опасность представляют органические сточные воды со специфическими токсическими свойствами, сбрасываемые кожевенными и целлюлозно-бумажными предприятиями. Они также вызывают дефицит кислорода за счет усиленного его потребления на биохимические и окислительные процессы.
Дефицит кислорода в загрязненном рыбохозяйственном водоеме может возникать вследствие угнетения фотосинтетических процессов в водоеме. Показательны в этом отношении загрязнения водоемов нефтью и нефтепродуктами.
Они образуют на поверхности водоема пленку, препятствующую нормальному газообмену между водой и атмосферой. Одна тонна нефти дает пленку в 10 км2.
Одновременно с этим нефтяная пленка затрудняет доступ солнечных лучей к фитопланктону, угнетая тем самым фотосинтез. Нефть и нефтепродукты подвергаются биохимическому окислению с интенсивным расходованием кислорода, ведущим к его дефициту в водоеме.
Нефть и нефтепродукты в количестве 15 мг/л абсолютно смертельны для всех живых существ, вызывая паралич дыхательных нервов.
Снижение фотосинтеза фитопланктоном и поверхностно-активные вещества (ПАВ), некоторые тяжелые металлы, многие пестициды.
Многие гербициды и альгициды оказывают угнетающее действие на процессы фотосинтеза низших и высших водных растений, снижая тем самым образование кислорода и его содержание в водоеме.
Нулевое содержание кислорода отмечается при поступлении гербицидов (монурона и диурона) в водоемы с большой биомассой макрофитов или при интенсивном развитии синезеленых водорослей. Массовое развитие синезеленых водорослей и их последующее отмирание и разложение, связанное с огромным расходом кислорода, также ведут к его дефициту в водоеме и могут быть одной из причин гибели рыб в цветущих водоемах.
Рыбам вреден и избыток кислорода. При перенасыщении воды газами, рыба также перенасыщается газами. При этом выделение газовых пузырьков происходит в тканях рыб. Пузырьки рвут кожу и плавники, выдавливают глаза, закупоривают кровеносные сосуды.
Перенасыщение воды кислородом наблюдается в водоемах при сильном освещении и мощном развитии зеленых водорослей.
Физиология дыхания рыб 2 страница
У костных рыб движения ротового и жаберного аппаратов сочетают нагнетательный (ротовая полость) и всасывающий (жаберная полость) принципы, что обеспечивает интенсивное продвижение воды сквозь жабры.
У акуловых рыб этот механизм выражен слабее: при быстром движении, сопряженном с большими затратами энергии, для покачивания воды через жабры используется само движение: рыба плывет с открытым ртом, и вода проталкивается через жабры тем интенсивнее, чем выше скорость движения. Такой же «пассивный» темп жаберной вентиляции отмечен у тунцов. При форсированном давлении это дает значительный (до 30%) энергетический выигрыш при критических скоростях.
Во всех случаях вода проходит сквозь жабры в одном направлении; движений типа «вдох-выдох» у водных позвоночных нет, вода прокачивается через жабры практически непрерывно давление ротовой полости на протяжении всего дыхательного цикла выше, чем в жаберной.
Извлечение О2 из воды в жабрах костистых рыб усиливается и вследствие того, что направление движения потока воды сквозь жаберные лепестки и тока крови в капиллярах, проходящих по вторичным жаберным пластикам, противоположны друг другу.
Такая противоточная система способствует тому, что на протяжении всей длины жаберной пластинки сохраняется градиент концентрации О2 (и СО2) в крови и в воде, благодаря чему процесс диффузии идет непрерывно, и отходящая от жабр кровь имеет почти тот же уровень насыщения О2, что и поступающая в жабры вода. Истинные жабры распологаются между двумя «насосами» – ротовым, который связан с работой мускулатуры дна ротовой полости и жаберным, который действует за счет движений жаберной крышки. В начале цикла рот открыт, и в обеих полостях существует небольшое отрицательное давление. Когда рот закрывается, давление в ротовой полости становится положительным, вода идет через жабры и в жаберной полости тоже возникает небольшое положительное давление. Когда жаберная крышка открывается, то давление в обеих полостях падает, в ротовой полости раньше, чем в жаберной; затем жаберная крышка закрывается, а рот открывается вновь.
У многих рыб обычный ритм дыхания время от времени прерывается: в результате закрытия жаберных крышек поток воды меняет направление на обратное и происходит «откашливание», очищающее жаберную полость.
При оценке эффективности дыхания нужно учитывать степень от сигенации крови, поступающей в жабры, по следующей формуле:
| Эффективность переноса = | Количество кислорода, переходящего из воды в кровь |
| Количество кислорода в крови при max насыщении |
или точнее
| Эффективность = | Vo2 | *100 |
| Qαво2[Pio2– Puo2] |
Vo2— поглощение кислорода
Q- объем крови, проходящей через жабры за данное время
αво2 — растворимость кислорода в воде (крови)
Pio2 — парциальное давление кислорода в протекающей воде
Puo2 — парциальное давление кислорода в венозной крови, входящей в жабры.
Максимальное поглощение кислорода достигалось бы в том случае, если бы оксигенированная кровь уравнивалась по величине Ро2 с приходящей к жабрам водой. Эффективность переноса О2 у форели приближается к 100%, однако рыба извлекает из воды лишь от 11 до 30% кислорода. Эта высокая эффективность указывает на то, что кровь течет навстречу воде. У поверхности жабр течение воды остается ламинарным, поэтому каждую жаберную пластику окружает тонкий не перемешивающийся слой, в котором кислород движется очень медленно. Сопротивление переносу кислорода в этом месте составляет 80-90% общего сопротивления при переходе кислорода из воды в кровь.
Гипоксия – недостаточность кислорода, как правило, вызывает у рыб рефлекторную брадикардию, т.е. увеличение ударного объема сердца, повышение сосудистого сопротивления в жаберном русле и на переферии, а также усиление гликолиза.
У рыб во время двигательной активности значительно снижается сопротивление в переферической сосудистой сети. При гипоксии эффективность поглощения кислорода снижается.
Подобные адаптации отмечены и в связи с кислородным режимом водоемов; содержание О2, отмечается удлинение жаберных лепестков, увеличение числа жаберных пластинок и возростание суммарной дыхательной поверхности, или увеличением объема воды, пропускаемой через жаберный аппарат. При гипероксии- повышении концентрации О2 дыхание, напротив, замедляется. Обычно реакция гипервентиляции сопровождается замедлением сердечного ритма (брадикардия), что отражает некоторое снижение уровня обмена веществ – метаболизма, соответственно и потребности в О2. Следовательно, «борьба за кислород» в условиях его временной недостаточности идет по двум путям: повышения интенсивности работы газообменного аппарата и некоторого снижения затрат О2 в организме. Чувствительность рыб к Ро2 значительно выше, чем к Рсо2. У форели уже при кратковременной гипоксии вентиляция возрастает в 7 раз.
При гипоксии некоторые рыбы могут использовать кислород содержащийся в газовом плавательном пузыре. У двоякодышащих рыб легкие отходят от брюшной стороны глотки; рыба дышит легкими во время длительного пребывания вне воды. Увеличение Рсо2 в воде вызывает остановку жаберного дыхания или снижает жаберное и стимулирует легочное дыхание.
4.1.3 Факторы, влияющие на потребление кислорода. Способность крови транспортировать кислород в связанном с гемоглобином состоянии зависит от общего количества гемоглобина (и соответственно от числа эритроцитов), а также от его химических свойств, в частности от сродства гемоглобина к кислороду, определяющего способность гемоглобина к насыщению кислородом при данном его парциальном давлении. У рыб, живущих в стоячих, богатых органикой водоемах, сродство гемоглобина к кислороду выше. Гематологические отличия связаны и с подвижностью рыб: обитатели хорошо аэрированных водоемов обычно высокоподвижны. Рыбы из водоемов с низким содержанием кислорода, обычно ведут малоподвижный образ жизни при появлении признаков кислородной недостаточности происходит выброс депонированных эритроцитов в кровь, что увеличивает ее кислородную емкость.
При повышении температуры вначале возрастает объем вентиляции, а затем и частоты дыхания, ритм сердца замедляется из-за недостаточного снабжения кислородом сердечной мышцы, и тогда работа обоих дыхательных механизмов может оказаться несогласованной.
Более активные рыбы имеют большую поверхность жабр, у окуня она почти в 2,5 раза больше, чем у камбалы. Противоток крови в капиллярах и омывающей жабры воды обеспечивает полное насыщение крови кислородом. При плавании рыбы ток воды может создаваться за счет движения с открытым ртом. Таким образом, жабры расположены как бы между двумя насосами создает прокачивание воды и вентиляцию жабр. За сутки через жабры прокачивается не меньше 1 м3 воды на 1 кг массы тела.
Внешнее дыхание. Под “внешним” дыханием, или газообменом понимают способы снабжения организма кислородом и удаление из него углекислоты, а под “тканевым” же дыханием понимают совокупность ферментативных реакций окислительных и неокислительных, в результате которых накапливается энергия в форме, доступной для биологической работы.
Интенсивность обменных процессов можно определить, измерить двумя путями: а) методом прямой калориметрии, б) непрямой калориметрии, а также по количеству потребляемого кислорода, образованного тепла или выделенной двуокиси углерода.
Концентрация кислорода падает с увеличением высоты. На дне прудов и озер, морских впадинах также наблюдается недостаток кислорода.
Концентрацию кислорода выражают по-разному, в воздухе газовую смесь выражают в % или парциальном давлении; жидкости – в миллиметрах, миллиграммах или молях растворенного газа на литр, в атмосферах – мм.рт.ст. парциального давления.
4.1.4 Способы дыхания. Существуют следующие способы дыхания: прямая диффузия, кожная, кишечное, легочное, дыхание через жабры, трахейное.
При нехватке кислорода в воде рыбы всплывают на поверхность воды и дышат атмосферным воздухом путем заглатывания. Электрический угорь в норме дышит воздухом через дополнительные дыхательные органы во рту, а также и кожным дыханием. Голец Misgurnus, живущий в прудах, при нехватке О², переходят от жаберного к кишечному дыханию – за счет заглатывания воздуха.
Прямая диффузия через поверхность кожи. Все поверхности клеток проницаемы для кислорода. При отсутствии транспортной системы, кислород проникает в организм рыб путем диффузии. Все многоклеточные животные в той или иной мере обладают кожным дыханием. У угрей через кожу поступает 60%, а через жабры всего лишь 40% кислорода. Водно — кожное дыхание относится к дополнительным приспособлениям, помогающим переносить неблагоприятные кислородные условия, т. е. использование растворенного в воде кислорода при помощи кожи, и воздушное дыхание—использование воздуха при помощи плавательного пузыря, кишечника или через специальные добавочные органы. Дыхание через кожу тела — одна из характерных особенностей водных животных. По интенсивности такого дыхания пресноводных рыб делят на три группы:
1. Рыбы, приспособившиеся жить в условиях сильного дефицита кислорода. Это рыбы, населяющие хорошо прогреваемые, с повышенным содержанием органических веществ водоемы, в которых часто наблюдается недостаток кислорода. У этих рыб доля кожного дыхания в общем дыхании составляет 17- 22%, у отдельных особей—42- 80%. Это карп, карась, сом, угорь, вьюн. При этом рыбы, у которых кожа имеет наибольшее значение в дыхании, лишены чешуи или она мелкая и не образует сплошного покрова.
2. Рыбы, испытывающие меньший недостаток кислорода и реже попадающие в неблагоприятные условия. К ним относятся рыбы, обитающие у дна, но в проточной воде, осетровые — стерлядь, осетр, севрюга. Интенсивность кожного дыхания у них составляет 9-12%.
3. Рыбы, не попадающие в условия дефицита кислорода, живущие в проточных или непроточных, но чистых, богатых кислородом водах. Интенсивность кожного дыхания не превышает 3,3 — 9%. Это сиги, корюшка, окунь, ерш. Через кожу происходит также выделение углекислоты. В извлечении кислорода из воздуха во влажной атмосфере участвует не только поверхность тела, но и жабры.
Кожное дыхание для представителей обитающих в водной среде имеет важное значение. Хорошо известен своим «наземным» образом жизни илистый прыгун Periophthalmus, обитающий в болотных эстуариях тропической зоны. Эта рыба подолгу находится вне воды, передвигаясь с помощью грудных плавников. Органом воздушного газообмена у нее служит кожа.
Важное значение при этом имеет температура. Наибольшей выживаемостью во влажной среде отличаются карась (11 сут), линь (7 сут), сазан (2 сут), в то же время лещ, красноперка, уклея могут жить без воды всего несколько часов и то при низкой температуре. При перевозке живой рыбы без воды кожное дыхание почти целиком удовлетворяет потребность организма в кислороде.
Жаберное дыхание. Основная масса водных позвоночных дышат через жабры. Жабры круглоротых и рыб обычно спрятаны, они вентилируются дыхательными движениями рта и жаберной крышки. У некоторых рыб и сами жабры обладают большой подвижностью.
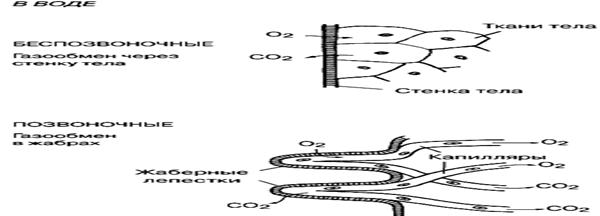 Рис. 35. Механизм газообмена у рыб А-через кожу; Б-через жабры
Рис. 35. Механизм газообмена у рыб А-через кожу; Б-через жабры
У двоякодышащей рыбы в гипоксической воде усиливается как легочное, так и жаберное дыхание. При повышении уровня СО2 в крови (гиперкапния) роль жабр в дыхании уменьшается, а дыхание воздухом усиливается. Хеморецепторы находятся на наружной стороне жабр или в выносящих жаберных сосудах. Гипоксия стимулирует вентиляцию, как легких, так и жабр; гиперкапния усиливает вентиляцию жабр в большей степени, чем легких, а при сочетании гипоксии с гиперкапнией, вентиляция легких возрастает, а жабр снижается.Использование жабр для воздушного дыхания возможно только при сохранении их поверхности во влажном состоянии.
Кишечное дыхание.У некоторых рыб, живущих в неблагоприятных условиях, выработались приспособления как, дыхание при помощи пищеварительного тракта — в различных отделах кишечника. В стенках кишечника образуются скопления капилляров. Воздух, заглатываемый ртом, проходит через кишечник, и в этих местах кровь поглощает кислород и выделяет двуокись углерода, при этом из воздуха поглощается до 50% кислорода. Такой вид дыхания свойствен вьюновым, некоторым сомовым и карповым рыбам; значение его у разных рыб неодинаково.
Воздушное дыхание рыб. Ряд видов костных рыб используют атмосферный воздух как дополнительный источник кислорода. Среди рыб с воздушным дыханием преобладают обитатели пресных вод или эстуариев тропической зоны. Данные отложения таких водоемов богаты органикой, интенсивное разложение ее при высокой температуре усиливает недостаток растворенного в воде кислорода.
Для воздушного дыхания могут использоваться жабры, слизистая ротовой и околожаберной полостей, кишечник, плавательный пузырь, кожа. Извлеченный из воды угорь вначале использует кислород, а затем начинает получать все больше кислорода через кожу, кроме того, дышит воздухом, проходящим через жабры. На воздухе жабры обеспечивают лишь одну треть, а в воде от 85 до 90% общего поглощения кислорода. При переходе из воды на воздух интенсивность метаболизма уменьшается вдвое и ритм сердца замедляется. У костистых рыб (линя, плотвы, форели) жабры расположены между ротовой и бронхиальной полостями: повышение давления в ротовой полости предшествует слабому насасыванию воды в бронхиальной полости. Этот насасывающий механизм хорошо развит у донных рыб. Такое чередование двух “насосов” имеется у акул и скатов.
Многие виды используют для воздушного дыхания плавательный пузырь, примеры такого типа газообмена – панцирная щука. У панцирной щуки путем воздушного дыхания в организм поступает 70-80% О2.
Настоящие легкие свойственны африканскому многоперу и двоякодышащим рыбам (Dipnoi), которые заселяют стоячие водоемы, подверженные регулярному пересыханию.
Другим способом использования атмосферного воздуха служит образование специальных добавочных органов дыхания как у лабиринтовых рыб, лабиринт — расширенный карманообразный участок жаберной полости, складчатые стенки которого пронизаны густой сетью капилляров, в которых происходит газообмен. Таким способом рыбы дышат кислородом атмосферы и могут находиться вне воды в течение нескольких дней (тропический окунь-ползун Anabas sp. выходит из воды и лазит по камням и деревьям).
4.1.5 Плавательный пузырь и газообмен Рыбы обладают специальными приспособлениями, чтобы держаться в толще воды. Основным органом, рыб регулирующих свой удельный вес, а также приуроченность к определенным слоям воды, является плавательный пузырь. Лишь немногие рыбы, обитающие в толще воды, его не имеют, это акулы и некоторых скумбриевых. Эти рыбы регулируют свое положение в том или ином слое воды только при помощи движения плавников.
Существуют два типа газовых пузырей: открытые у открытопузырных рыб и закрытые у закрытопузырных. Открытопузырные могут заглатывать в пузырь воздух; кроме того, и те и другие рыбы обладают способностью выделять в полость пузыря газ. Газовый (плавательный) пузырь выполняет в первую очередь гидростатическую роль, и регулируют глубину погружения. Так глубоководные рыбы всплывают или погружаются в зависимости от объема газа в пузыре.
Вторая функция плавательного пузыря – повышение чувствительности сенсорных органов: пузырь усиливает звуковые волны, передаваемые водой. У некоторых рыб плавательный пузырь служит для испускания звуков. Кроме того, плавательный пузырь может играть роль поставщика кислорода при респираторном стрессе, как, например, у данных рыб. Плавательные пузыри более широко распространены у поверхностных и пелагических рыб, обитающих на глубине до 600 м., чем у донных.
Плавательный пузырь служит не только для изменения удельного веса рыбы, он играет роль и органа, определяющего величину наружного давления у ряда рыб. У большинства вьюновых – Cobitidae, ведущих данный образ жизни, плавательный пузырь сильно редуцирован, и его функция как органа, воспринимающего изменения давления, является основной. Рыбы могут воспринимать даже незначительные изменения давления, их поведение меняется при перемене атмосферного давления. В Японии некоторых рыб специально содержат для этой цели в аквариумах и по изменению их поведения судят о предстоящей смене погоды.
Для поддержания плавучести в морской воде нужен плавательный пузырь, объем которого составлял бы 3-5% объема тела, а в пресной воде — несколько больше. Закрытопузырные рыбы наполняют пузырь газом исключительно путем секреции, открытопузырные же могут заглатывать часть газа, но при отсутствии доступа к поверхности воды они способны секретировать его. Плавучесть рыб, лишенных плавательного пузыря, часто достигается за счет увеличения собственного веса, в частности жирности. В особенности увеличивается вес печени, что составляет 23% от веса тела, а ее удельный вес- 0,94.
У большинства более древних групп рыб плавательный пузырь соединен с кишечником при помощи специального протока. У остальных рыб – окунеобразных, трескообразных и других костистых, с возрастом связь плавательного пузыря с кишечником не сохраняется. Плавательные пузыри у некоторых сельдевых и анчоусов, у океанической сельди, шпроты, хамсы имеют два отверстия. У рыб, имеющих соединение плавательного пузыря с кишечником, поступление и выделение газа из плавательного пузыря происходит в значительной степени через ductus pneumaticus, это наружное отверстие в задней части пузыря, открывающееся непосредственно за анальным отверстием. Это отверстие позволяет рыбе за короткий срок удалить из плавательного пузыря лишний газ. При этом у опускающейся на глубину рыбы излишний газ появляется в пузыре под воздействием возрастающего по мере погружения рыбы давления воды на ее организм.
За исключением некоторых сельдевых все рыбы, обладающие плавательным пузырем, не могут быстро переходить из поверхностных слоев на глубины и обратно, а закрытопузырные рыбы могут перемешаться в слое воды. В связи с этим у большинства видов, совершающих быстрые вертикальные перемещения, плавательный пузырь либо совершенно отсутствует, либо редуцирован, а удержание в толще воды осуществляется за счет мускульных движений тела или плавников. Редуцируется плавательный пузырь и у многих донных рыб, многих бычков, морских собачек, вьюновых и др. Редукция пузыря у донных рыб, связана с необходимостью обеспечить больший удельный вес тела.
Наполнение газом плавательного пузыря происходит не сразу по выходе из икринки. Пока выведшиеся свободные эмбрионы проходят стадию покоя, подвесившись к стеблям растений или лежа на дне, газа в плавательном пузыре у них нет. Наполнение плавательного пузыря газом у одних видов осуществляется за счет наружного воздуха, у других газ выделяется крупными клетками, расположенными внутри плавательного пузыря. У многих рыб проток, соединяющий кишечник с пузырем, во взрослом состоянии отсутствует, а у их личинок он имеется.
Секреция газа в плавательный пузырь. Некоторые водные животные способны накачивать газы в пузырях или газовых камерах против значительного градиента давления. Наиболее изучен механизм данного процесса секреции газов в плавательный пузырь у костистых рыб. У закрытопузырных рыб весь газ плавательного пузыря происходит из газовой железы. Наибольшее количество газа секретируется у открытопузырных.
У большинства рыб, обладающих плавательным пузырем, имеются механизмы, как для секреции газа, так и для его поглощения. В регулировании объема пузыря могут участвовать железы и мышцы. Мышцы, ответственные за наполнение пузыря воздухом, снабжены нервными холинэргическими, а их антагонисты – адренэргическими окончаниями. Существующие медиаторы действуют как на мышцы, так и на железы.
У рыб с замкнутым плавательным пузырем после первоначального наполнения газом извне дальнейшее изменение количества и состава газа регулируется путем его выделения и поглощения кровью. У таких рыб на внутренней стенке пузыря находится красное тело – образование, чрезвычайно густо пронизанное кровеносными капиллярами. Так, в двух красных телах плавательного пузыря угря имеется 88000 венозных и 116000 артериальных капилляров (в красных) общей длиной 352 и 464м. В то же время объем всех капилляров в красных телах угря составляет всего лишь 64мм 3. По величине красное тело варьирует у рыб от небольшого пятна до мощной газоотделительной железы, состоящей из цилиндрического железистого эпителия. Скорость наполнения газом плавательного пузыря за счет газов крови различна, так рыбе с открытым пузырем для наполнения плавательного пузыря требуется 5-7 дней, а угрю 12-20 ч.
Газ, выделяемый в плавательный пузырь, сильно отличается по составу от воздуха. На глубине, где действует гидростатическое давление, количество растворенного газа возрастает на 14% при каждом увеличении давления на 100 атм., так что на глубине 1000 м, Ро2 достигает 0,228 атм. Газ в плавательном пузыре глубоководной рыбы может содержать 65-95% О2. Таким образом, общее давление кислорода в плавательном пузыре рыбы на глубине 1000 м составляет около 90 атм, т.е. активный перенос кислорода в плавательный пузырь увеличивает давление О2 в 394 раза.
Состав газа в плавательном пузыре у морских рыб, взятых на глубине 100 м составил: так, количество азота соответствует 0,8% давления воздуха, или 0,8 атмосферного давления, на глубине 900 м., парциальное давление азота в плавательном пузыре колебалось от 5 до 15 атм. Когда на поверхности воды напряжение азота соответствует 80% давления воздуха — 0,8 атмосфер. На каждые 10м. глубины давление повышается приблизительно на 1 атм. Следовательно, у рыбы на глубине 100 м. при выделение газа должен преодалеваться градиент давления в 10 атм. Если рыба всплывает, расширение газа может вызвать разрыв плавательного пузыря.
Состав газа в плавательном пузыре неодинаков, как у различных видов рыб, так и у разных особей одного и того же вида. В плавательном пузыре морских рыб кислорода содержится больше, чем у пресноводных.
Каждому виду свойствен свой кислородный порог, т. е. минимальная концентрация кислорода, при которой рыба гибнет. Форель начинает задыхаться при концентрации кислорода 1,9 мг/л, судак и лещ погибают при 1,2, плотва и красноперка — при 0,25 — 0,3 мг/л; у сеголетков-карпов, выращенных на естественной пище, кислородный порог отмечен при 0,07- 0,25 мг/л, а для двухлетков—0,01-0,03 мг/л кислорода. Караси и ротаны — частичные анаэробы — несколько суток могут жить совсем без кислорода, но при низкой температуре. Предполагают, что сначала организм использует кислород из плавательного пузыря, затем — гликоген печени и мышц.
Подвижные и хищные рыбы имеют большой запас кислорода в плавательном пузыре, который расходуется организмом при бросках за добычей, когда поступление кислорода через органы дыхания оказывается недостаточным. В неблагоприятных кислородных условиях воздух плавательного пузыря у рыб используется для дыхания. Вне воды угорь использует для дыхания кислород плавательного пузыря и воздух, проходящий через кожу и жабры. Это позволяет ему даже переползать из одного водоема в другой. Осваивая различные водоемы, рыбы приспособились к жизни при разных газовых режимах. Наиболее требовательны к содержанию кислорода в воде лососевые, которым для нормальной жизнедеятельности нужна концентрация кислорода 4,4-7 мг/л; хариус, голавль, налим хорошо себя чувствуют при содержании не менее 3,1 мг/л; карповым обычно достаточно 1,9- 2,5 мг/л.
На рыб губительно действует чрезмерное перенасыщение кислородом воды. Так, летальной границей для эмбрионов щуки является 400% насыщения воды кислородом, при 350 — 430% насыщения нарушается двигательная активность эмбрионов плотвы. Прирост осетровых снижается при 430% насыщении кислородом.
Инкубация икры в перенасыщенной кислородом воде приводит к замедлению развития эмбрионов, сильному увеличению отхода и количества уродов и даже гибели. У рыб появляются пузырьки газа на жабрах, под кожей, в кровеносных сосудах, органах, а затем наступают судороги и смерть. Это называется газовая эмболия или газопузырьковая болезнь. Для поддержания оптимальной концентрации кислорода в воде, обеспечивающей наиболее эффективное течение физиологических процессов в организме рыб, нужно использовать аэрационные установки. У них повышается обмен и как результат увеличивается потребление корма и снижается кормовой коэффициент, развитие эмбрионов ускоряется, отходы снижаются.
Наибольшего развития как орган дыхания плавательный пузырь достигает у двоякодышащих рыб. У них он ячеистый и функционирует, как легкое. При этом возникает “легочный круг” кровообращения. Благодаря переплетению сосудов со встречными потоками крови кислород может переходить из вены, идущий от плавательного пузыря, непосредственно в артерию, идущую от сердца, а затем из общей вены в артерию плавательного пузыря. Таким образом, в ткани секретирующей железы может создаваться высокая концентрация кислорода. В артерии плавательного пузыря имеет место «положительный» эффект Рута, т.е. увеличение максимального уровня насыщения гемоглобина.
У некоторых глубоководных рыб полость плавательного пузыря содержит большое количество липидного материала, который состоит из фосфолипида и холестерина в соотношении 1:1. Газовая железа глубоководных рыб синтезирует холестерин. Так как кислород, возможно, хорошо растворим в этом холестеринсодержащим липиде, в пузыре может оставаться очень мало свободного газа. Напротив, у рыб, обитающих на небольших глубинах, имеется лишь тонкий слой липида на внутренней поверхности пузыря.
Сифонофоры как физалия выделяют газ в объемистый пузырь-пневмотофор. Секретируя газ, она всплывает на поверхность, а выпуская его – погружается под воду. Находящийся в пузыре газ содержит 74,4% N2, 8,9% СО, 14,4% О2, 1,1% аргона и 0,4% СО2. У основания пузыря расположена газовая железа, сходная по своей ультраструктуре с железой рыб. Формы у этой сети и железы у разных рыб разная, но принцип работы одинаковый; артерии и вены в железе распадаются на множество капилляров, а затем вновь собираются в один сосуд. Капилляры по длине достигают до 25 мм, тогда как в мышцах длина равна 0,5 мм. Газовая железа в пищеварительном пузыре выполняет также роль умножителя благодаря своему морфологическому строению, именно здесь, наблюдается высокое парциальное давление газов. У глубоководных рыб благодаря чудесной сети количество кислорода находится на нужном уровне из-за его непрерывного и диффузного физического поступления в артериальные капиляры.
Интенсивность дыхания рыб зависит от размера, возраста, подвижности, активности питания, пола, степени зрелости гонад, физико-химических факторов среды. По мере роста рыб активность окислительных процессов в тканях уменьшается, а созревание гонад, наоборот, вызывает увеличение потребления кислорода. Расход кислорода в организме самцов выше, чем у самок.
4.1.6.Регуляция процессов дыхания. Физиологические механизмы регуляции ритма дыхания у рыб изучены еще недостаточно. Ритм дыхательных движений поддерживается импульсами, идущими от клеток дыхательного центра, расположенных в продолговатом мозге. Ритмические разряды нервных импульсов в этой области можно обнаружить даже после обездвиживания рыбы тубокурарином или сукцинилхолином, а также после перерезки всех чувствительных нервных волокон, идущих к этой части мозга. Локализация кислородных рецепторов точно не выяснена. Скорее всего, они расположены не в самих жабрах (брадикардия возникает и при отключении тока крови через жабры), а на их поверхности, в спинной аорте, в ротовой полости или же в ложножабрах.
У рыб есть как периферические, так и центральные рецепторы воспринимающие уровень О2. Недостаток кислорода увеличивает как частоту, так и глубину дыхания, поэтому объем вентиляции возрастает. Избыток СО2 иногда может вызывать небольшое снижение частоты и углубление дыхания. Угорь Anguilla и форель Salmo поглощают около 80% — О2, растворенного в воде, насыщенный воздухом; при более низком содержании О2 в воде они поглощают меньше кислорода. Если уровень О2 уменьшается до 4 мл/л., то угорь и форель усиливают в 5 раз интенсивность дыхательных движений. Четыре – пяти кратное увеличение активности приводит к тому, что поглощение О2 из воды повышается на 70%, в результате рефлекторного воздействия накопившегося углекислоты.
Наиболее тонкая регуляция дыхательного процесса и работы дыхательного центра, расположенного в продолговатом мозге осуществляется за счет концентрации углекислоты в крови. Углекислота является крепким стимулятором дыхательного центра, обладает высокой растворимостью в воде и редко накапливается в тканях водных животных при нарушение их дыхания. Первичным возбудителем центра дыхания является недостаток О2 в воде. Дыхательный центр акульих и костных рыб, хорошо развиты, обладают автономной активностью. У акулы во всех жаберных ветвях девятого и десятого черепномозговых нервов имеются ингибиторные эфферентные волокна. У сома и карася центры регуляции находятся позади моторного ядра, у линя в нижнем краю фасциальных долей продолговатого мозга.
§
Контрольные вопросы.
1.Каковы особенности строения и основные функции жаберного аппарата рыб?
2.Типы дыхания у рыб.
3.Каков механизм вдоха и выдоха у рыб?
4.Перечислите основные формы транспорта кислорода и углекислого газа кровью.
5.Охарактеризуйте понятия “внешнее”и “тканевое” дыхание или газообмен.
6.Механизм нервной и гуморальной регуляции процессов дыхания.
7.Чем характеризуется кислородный порог у рыб?
8.Морфологические и физиологические особенности плавательного пузыря у различных видов рыб?
9.Какова роль плавательного пузыря в жизнедеятельности рыб?
10.Каков газовый состав воздуха плавательного пузыря?
ГЛАВА 5
§
5.1Способы питания рыб.Способы питания зависят от типов пищи, среди которых можно выделить такие: 1) органические молекулы, растворенные в тканях или естественных водах; 2) мелкие частицы, в основном мертвый детрит, а частично- живой фитопланктон; 3) крупные частицы, куски пищи или пойманная жертва.
Вода в естественных водоемах содержит органические вещества в концентрации 2-20 мг/л. В океанической воде эти показатели сильно изменяются в зависимости от места и сезона. Растворенные органические молекулы активно поглощаются некоторыми водными животными. Хотя поглощаемых таким образом количеств недостаточно для жизнедеятельности организма, тем не менее, они могут вносить существенный вклад в питание этих животных.
По характеру питания рыб внутренних водоемов делят:
Растительноядные — в пищу употребляют в основном водную флору (белый амур, толстолобик).
Животноядные — которые употребляют в пищу беспозвоночных (лещ, вобла и др.). Они делятся на две группы: Планктофаги — питаются водорослями (фитопланктон), некоторыми кишечноподобными, моллюсками, яйцами и личинками беспозвоночных и др: Бентофаги — питаются организмами, обитающими на грунте и в грунте дна водоема:
Ихтофаги, или хищные, которые питаются рыбой, позвоночными животными (лягушки, водоплавательные птицы). Однако такое деление условно. Многие рыбы имеют смешанное питание. Например, карась — всеяден.
При любом типе питания в организм попадают строго определенные группы веществ: вода, минеральные соли и органические вещества, из которых наиболее важны белки, жиры и углеводы. Вода и минеральные соли легко всасываются в пищеварительном тракте и поступают в кровь без какой-либо предварительной перестройки
Рыбы питаются строго по графику, сменяя друг друга в течение дня не только по видовому признаку, но и по возрастным группам. Графики питания меняются посезонно, рыбы питаются в течение всего дня, некоторые дневные рыбы переходят на питание в сумеречное, и в ночное время, сроки питания рыб четко укладываются в строгие временные рамки.
Лещ в мае потребляет около 35% годового количества пищи, в июне — около 30%. Язь в июле съедает 25% пищи, в августе — 20%. Сом в мае-июне потребляет 75% годового рациона. Окунь в мае-июне съедает 30-35%, а в сентябре — 20% годового рациона. Рыбы легко «переедают», накапливают про запас энергию в виде жира, весьма насыщенный витаминами и макро и микроэлементами. Поэтому, рыбы и способны продолжительное время пережидать погоду.
Рыб, делят по этому признаку на «диетических» и мирных». К первым относят судака, окуня, ко вторым — сазана, сома, линя и других.
 Диетические рыбы не способны к зимней по перволедью спячке, у некоторых из них в течение года наблюдается запланированный жор 3-4 кратности; 1) короткий преднерестовый жор – послезимний — летний период; 2) посленерестовый — связан со временем восхода и захода;3) предзимний жор и жор, наступающий на третий- четвертый день после становления льда.
Диетические рыбы не способны к зимней по перволедью спячке, у некоторых из них в течение года наблюдается запланированный жор 3-4 кратности; 1) короткий преднерестовый жор – послезимний — летний период; 2) посленерестовый — связан со временем восхода и захода;3) предзимний жор и жор, наступающий на третий- четвертый день после становления льда.
Питание хищной рыбы протекает периодически, например щука при полном желудке переваривает пищу в течение 1—3 суток. В это время интенсивность питания, конечно, понижается. При недостаточности корма рыба питается в течение всех суток. Вся жизнь рыб протекает в полной гармонии со временем: четко расписана и по времени суток, и по сезонам.
Рыб по образу жизни во времени делят на дневных, сумеречных, ночных. Все физиологические функции четко привязаны к определенным периодам сезона. Рыбы питаются в течение суток по четкому графику, который также четко меняется с изменением сезонов и т.д. Сезонное поведение в отличие от суточного определяется действием не только световогофактора, так как их биологические часы настроены на поток света, исходящий от Солнца, сила которого меняется и течение суток, и посезонно, что приводит к изменению и поведения рыб.
От светового потока зависит и температура воды, которая активизирует поведение и питание рыбы. Например, сом в мае-июне съедает 75% годового рациона своей пищи, ему в этот период для насыщения ночного времени не хватает, он начинает питаться и днем, т.е. становится уже не ночным и даже не сумеречным животным. Установлено, что у рыб светочувствительные клетки разбросаны по всей поверхности тела которые формируют у рыб «третий глаз» не имеющий отношения к зрению, но вызывают реакцию на световой поток.
Суточное питание рыб связан со временем восхода и захода солнца, и рыбы четко придерживаются этих ориентиров, но они в определении времени могут ошибаться в пределах 10 минут и то только в редких случаях. Сезонные и годовые ритмы в наших широтах зависят от температурного фактора. Тело рыбы в спокойном состоянии имеет температуру, равную температуре окружающей воды. Рыбы очень чувствительны к ее изменению, различая разницу в 0,1°С. Чем выше температура воды (до известных пределов), тем переваривание пищи у рыбы протекает быстрее и, следовательно, рыба должна принимать корм чаще.
В жаркую летнюю погоду рыба обычно кормится меньше, весной и ранней осенью — больше. Следует отметить, что температура воды в питании рыбы играет важную роль, каждая порода рыб принимает наибольшее количество корма только лишь при самой благоприятной для нее температуре. Например, наиболее интенсивное питание щуки происходит при температуре воды около 12—18° С. При понижении температуры воды до 5—7° С., жизненные функции рыбы замедляются, следовательно, в какой-то степени понижается и интенсивность питания.
5.2 Взвесь питательных веществ.Такую взвесь составляют частицы детрита, органические соединения, адсорбированные на инертных частицах, мертвый планктон, а также живой фитопланктон – диатоновые водоросли, жгутиковые и бактерии. Количество фитопланктона зависит от времени года, географического положения и глубины; в прибрежных морских водах его содержание доходит до 700 мкг/л. Пресные озера тоже могут быть богаты фитопланктоном (2 мг/л), особенно летом в поверхностном слое, зимой он может исчезать. Многие животные питаются только такой суспензией, прокачивая большие количества воды через свои фильтрующие органы. Существуют два основных способа питания путем фильтрации – использование либо слизистых сетей, либо ресничек и жгутиков.
Макрокомпоненты пищи включают углеводы, белки, жиры и нуклеиновые кислоты. В ходе эволюции в желудочно – кишечном тракте рыб сформировался набор пищеварительных ферментов, который способен расщеплять каждое из перечисленных выше сложных органических веществ до простых единиц, всасываемых и включаемых затем в обмен веществ.
5.3 Пищеварение

 .Все животные в качестве основных питательных веществ используют набор определяемых макромолекул. Для того чтобы эти вещества могли участвовать в метаболизме, необходимо расщепить их до простых форм с помощью специальных катализаторов — ферментов. Под термином «пищеварение» обычно подразумевается процессы расщепления сложных органических и неорганических веществ, в более простые и легкоусвояемые формы.
.Все животные в качестве основных питательных веществ используют набор определяемых макромолекул. Для того чтобы эти вещества могли участвовать в метаболизме, необходимо расщепить их до простых форм с помощью специальных катализаторов — ферментов. Под термином «пищеварение» обычно подразумевается процессы расщепления сложных органических и неорганических веществ, в более простые и легкоусвояемые формы.
Ферменты, участвующие в пищеварении, относятся исключительно к гидростатическому типу. Дополнительным субстратом, участвующим в реакциях в пищеварительном тракте, является вода. Пищеварительная система позвоночных представляет собой своего рода «конвейер», пища поэтапно претерпевает механические и химические изменения, в результате которых конечные легкоусвояемые продукты пищеварения проникают сквозь стенку кишечника в кровяное русло.
Пищеварительный тракт – это трубка, представленная многочиленными сложными вариантами, приспособленными к типу используемой пищи и способу питания организма. У рыб отмечается следующая последовательность питания:
А) Прием пищи. Ротовое отверстие и его придатки отражают все разнообразие форм в разных группах животных, которое обусловлено особенностями способов поглощения пищи. В процессе используются зубы, для смазывания пищи слизью.
Б) Начальное переваривание. У некоторых рыб имеются структуры для внутреннего растирания или перемешивания пищи (терки, «желудочные мышцы», мускульный желудок), отделы желудка, выполняющие механическую функцию- МОТОРИКУ. (Рис. 36)
Рис.37. Моторика желудка щуки (а) и кишечника карпа (б)
Средняя кишка или желудок, является основным местом переваривания пищи. Пищеварительные ферменты вырабатываются здесь слизистой желудка, либо железистым выростом, или дивертикулом, гепатопанкресан. Эта железа иногда несет чисто секреторную функцию. У акуловых рыб экзокринная часть поджелудочной железы представлена ацинарными клетками и функционально полностью дифференцирована, как и у всех высших позвоночных.
В) Завершение переваривания и всасывания питательных веществ. Эти процессы идут либо в большей части кишечника, либо в задней части средней кишки. Кишечник имеет сложное строение; его поверхность образует многочисленные складки. Ферменты вырабатываются щеточной поверхностью эпителия. К таким ферментам относятся – карбогидразы, липазы, энтеркиназы, некоторые пептидазы и щелочная фосфатаза. В конечной стадии пищеварения, вода обратно всасывается в организм и идет процесс формирования каловых масс.
Пища поступает в организм через ротовое отверстие. Ротовой аппарат предназначен для захвата пищи; если пищевые объекты крупные, то отчленяются куски, доступные для проглатывания.
У многих животных пища частично механически обрабатывается в ротовой полости (раздавливание, расчленение, надрезание). Измельчение пищи способствует ее более эффективному перевариванию. Расположенные в ротовой полости железы и их жидкий секрет, увлажняют и обволакивают пищевые комки, что облегчает их передвижение по пищеводу.
Приспособления, связанные с захватом пищи, включают челюстной аппарат, зубную систему и жаберноглоточный аппарат.
Черты строения ротового аппарата, обусловленные характером питания и способом захвата пищевых объектов. Форма рта у рыб разнообразна. Рыбы, питающиеся планктоном, имеют верхний рот, питающиеся у дна — нижний рот, у хищных рыб – конечный рот. За ртом у рыб находится ротовая полость, куда первоначально поступает пища.
Расположение рта на голове определяются расположением пищевых объектов относительно рыбы. У видов, приспособленных к захвату пищевых объектов, находящихся спереди или же в самых различных положениях, рот расположен на конце головы – конечный и обращен вниз (Acepenseridae, Gobio, Borbus, Chond rostoma, Mullus, Mugil и т.п.).
Рис.38. Различные формы рта: а – верхний; б – конечный; в – полунижний; г – нижний; д – выдвижной; е — воронкообразный (круглый)
Когда пищевые объекты находятся сверху, рот становится обычно верхним, т.е. располагается на верхней стороне головы (Esox, Pelecus, Silecus, Siranoscopus, Lophins и др.). Рис.37.
Будучи в закрытом состоянии верхний, а в открытом состоянии нижняя часть рта в момент действия, такой рот становится конечным, так как при открытом состоянии ротовое отверстие направлено прямо вперед.
Относительная величина рта у рыб определяется рядом факторов и прежде всего зависит от размеров пищевых объектов. Относительные размеры рта зависят и от плотности распределения пищевых объектов в толще воды. Чем ниже плотность распределения пищевых объектов, тем больше относительные размеры рта.
У рыбоядных видов, питающихся сравнительно крупными подвижными объектами величина рта зависит от способа поимки жертвы.
У хищников преследующего типа (Scombridas, Carangidae, Xiphiidae) рот, как правило, относительно меньше, чем у хищников подстерегающего типа. Догоняющий хищник ловит жертву своей скоростью и поворотливостью, тогда как подстерегающий хищник делает только один бросок на жертву и вероятность ее поимки зависит, от относительных размеров рта, увеличение которого может компенсировать неточность броска.
При любом расположении и относительной величине рот может быть невыдвижным или выдвижным.
Невыдвижной рот характеризуется неподвижным или почти неподвижным соединением середины верхней челюсти с черепом. Рот этого типа свойственен, как правило, рыбам, питающимся сравнительно крупными объектами и в процессе захватывания пищи вынужденным затрачивать значительные усилия на закрывание рта. Это характерно, прежде всего, для большинства рыбоядных форм, которым приходится с заметным усилием сводить челюсти для удержания схваченной жертвы (Esocidae, большинство Scombridae и др.), а также для многих бентофагов разрывающих раковины моллюсков, твердые панцири ракообразных, иглокожных.
Выдвижной рот характеризуется подвижным соединением середины верхней челюсти с черепом, поэтому при раскрывании рта характерен преимущественно для рыб, питающихся сравнительно мелкими объектами: планктоном.
Обыкновенно различают несколько типов рта у рыб: хватательный, всасывающий, дробящий, рот планктоноеда, рот перифитопоеда. Все эти типы, связанные с характером питания, отличаются не только внешней особенностью рта, но и внутренными, анотомическими чертами (строением отдельных элементов скелета ротового аппарата, мускулов, особенностями зубной системы и др.).
Хватательный и всасывающий выполняют хватательную функцию (громадное большинство рыб), а всасывающий, характеризуется практически полной утратой этой функций челюстей. Выделены случай полной функциональной атрофии ротового аппарата связанные с паразитическим образом жизни.
У хрящевых рыб как акулы рот снабжен подвижными хрящевыми челюстями. На коже, покрывающей челюсти, плакоидные чешуи преобразовались в крупные конические, загнутые вершинами назад зубы, которые располагаются несколькими рядами. На дне ротовой полости находятся язык как небольшая складка слизистой оболочки, поддерживаемая непарным элементом подъязычной дуги, собственной мускулатуры он не имеет. В ротовой полости костистой рыб находятся острые, слегка направленные назад конические зубы.
5.4 Пищеварительная система хрящевых рыб.В пищеварительном тракте хрящевых рыб различают; ротовую полость, глотку, пищевод, желудок, кишечник (тонкая, толстая, прямая кишка, заканчивающаяся анусом). У акул, скатов и некоторых других рыб перед анусом имеется клоака — расширение, куда открывается прямая кишка и протоки мочевой и половой систем.
Ротовая полость без заметной границы переходит в обширную глотку, пронизанную жаберными щелями. Выпадению пищи через жаберные щели препятствуют сидящие на жаберных дугах палочковидные хрящи-жаберные тычинки.
От задней части глотки начинается пищевод (oesophagus). Пищевод без заметных границ переходит в обширный, легко растяжимый U-образный желудок (gaster). От желудка отходят короткая тонкая кишка (collum). За ней следует прямая кишка (rectum), которая открывается в клоаку. Внутри толстой кишки имеется просвечивающий сквозь ее стенки так называемый спиральный клапан: он представляет собой складку слизистой оболочки, которая вдается в просвет кишки. Складка расположена спирально: образует около десятка витков. Рис. (39)
Рис.39. Пищеварительная система акулы

Спиральный клапан значительно увеличивает всасывающую поверхность. От средней части прямой кишки отходят полый пальцеобразный вырост — ректальная железа (glandula rectalis) –орган солевого обмена. В период размножения ректальная железа выделяет пахучую слизь, которая в виде тонких нитей остается позади плывущей рыбы. Это облегчает встречу особей разного пола.
Железы пищеварительного тракта у акуловых рыб хорошо развиты. Массивная трехлопастая печень (hepar) занимает переднюю часть брюшной полости. Вырабатываемая в печени желчь собирается в желчный пузырь (vesia fellea), связанный с тонкой кишкой отчетливо заметным желчным протоком. Компактная поджелудочная железа (pancreas) прилегает к тонкой кишке, сообщаясь с ней мелкими протоками. К изогнутой части желудка примыкает компактная, с заостренным краем селезенка (lien) – орган кроветворения.
5.5 Пищеварительная система костной рыбы.Без четких границ ротовая полость переходит в глотку, прободенную жаберными щелями. В глубине глотки начинается короткий пищевод, который почти сразу переходит в желудок (gaster). За желудком следует кишечник, слабо дифференцированный на двенадцатиперстную (duodenum), тонкую (collum), и прямую (reetum) кишки. Прямая кишка открывается наружу анальным отверстием. Сразу за сердцем в передней части брюшной полости под желудком располагается крупная печень (hepar). На ее внутренней стороне находится желчный пузырь (vesica fellea) –полость, в которой скапливается вырабатываемая в печени желчь. От желчного пузыря начинается желчный проток (ductus choledochus), впадающий в начало двенадцатиперстной кишки. Вдоль желчного протока лежит поджелудочная железа (pancreas). В месте перехода желудка в двенадцатиперстную кишку (первый изгиб кишечника) рядом с желудком расположена компактная селезенка (lien).

Рис.40. Внешнее строение окуня:
1-почки; 2 – плавательный пузырь;
3 – половая железа;
4 – мочевой пузырь;
5 – кишечник; 6 – желудок;
7 – пилорические придатки;
8 – печень
9 – пищевод; 10 – сердце
11 – жабры; 12 – селезенка
Над кишечником в верхней части брюшной полости находится крупный плавательный пузырь (vesica natatoria), который служит гидростатическим органом. Плавательный пузырь щуки узким протоком связан с передней частью кишечника. У многих других рыб (например, карповых, окуня и др.) во взрослом состоянии плавательный пузырь полностью изолирован от кишечника.
В ротовой полости рыб нет слюнных желез. Железистые клетки ротовой полости и глотки выделяют слизь, которая не имеет пищеварительных ферментов и способствует лишь проглатыванию пищи, а также защищает эпителий ротовой полости с вкрапленными вкусовыми почками (рецепторами). Мощный и выдвижной язык имеют только круглоротые, у костистых рыб он не обладает собственной мускулатурой.
Рот обычно снабжен зубами. Наличием эмалевого колпачка и слоем дентина они напоминают зубы высших позвоночных. У хищников они располагаются как на челюстях, так и на других костях полости рта, иногда даже на языке; они острые, часто крючкообразные, наклонены внутрь к глотке и служат для схватывания и удержания жертвы. У многих мирных рыб (многие сельдевые, карповые и др.) на челюстях зубов нет.
5.6 Механизм питания координирован с дыхательным механизмом. Вода, всасываемая в рот при вдохе, несет и мелкие планктонные организмы, которые при выталкивании воды из жаберной полости (выдох) задерживаются в ней жаберными тычинками. Они так тонки, длинны и многочисленны у рыб, питающихся планктоном (планктонофагов), что образуют фильтрующий аппарат. Отцеженный комочек пищи направляется в пищевод. Хищные рыбы не нуждаются в отфильтровывании пищи, тычинки у них редкие, низкие, грубые, острые или крючковатые: они участвуют в удержании жертвы. У некоторых бентосоядных рыб на задней жаберной дуге имеются широкие и массивные глоточные зубы, они служат для перетирания пищи.
Строение, форма и длина пищеварительного тракта разнообразны в связи с характером пищи (объектами питания, их усвояемостью), особенностями переваривания. Рис.40.
Имеется определенная зависимость длины пищеварительного тракта от рода пищи. Так, относительная длина кишечника (отношение длины кишечника к длине тела.) составляет у растительноядных (пинагора и толстолобика)- б-15, у всеядных (карася и карпа) — 2- 3, у хищных (щуки, судака, окуня) — 0,6-1,2.
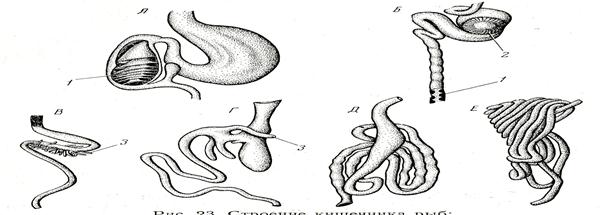
Рис.41. Строение кишечника рыб:
А – скат; Б – осетр;В – лосось; Г – окунь; Д – карп; Е – толстолобик;
1 – спиральный клапан; 2 – пилорическая железа;3 – пилорические придатки
Следующий за глоткой пищевод проводит пищу в желудок, обычно короткий, широкий и прямой с сильными мускулистыми стенками. В стенках пищевода имеются многочисленные клетки, выделяющие слизь. У открытопузырных рыб в пищевод открывается проток плавательного пузыря.
Желудок представляет собой первый отдел пищеварительной системы, в котором начинается ферментативное расщепление пищи, в частности расщепление белков в кислой среде. В слизистой оболочке желудка имеются железистые клетки, вырабатывающие желудочный сок, в составе которого имеется соляная кислота и пепсин, расщепляющий белок в кислой среде, создающую нужную РН реакцию среды и слизь. Здесь у хищных рыб переваривается основная часть пищи.
Форма и строение желудка сильно различаются в разных группах животных. У некоторых «мирных» рыб желудок вообще морфологически не выражен, хотя функционально существует. У других рыб, также он представляет собой определенную форму расширения отдела кишечной трубки. Железы, расположенные на дне желудка, выделяют щелочной и кислый секрет, содержащий соляную кислоту. Не все рыбы имеют желудок. Он отсутствует у карповых, многих бычков и некоторых других рыб.
Из желудка пища отдельными порциями, в разжиженном состоянии в виде химуса поступает в кишечник. В кишечнике осуществляется ферментативное воздействие на пищу при щелочной реакции среды, а также всасывание конечных продуктов расщепления. Начальный отдел тонкого кишечника осуществляет наиболее активные процессы с участием ферментов поджелудочной железы и желчи, попадающий сюда из печени, железы, расположенные в стенках кишечника-люберкюновые, также выделяют пищеварительный кишечный сок богатый ферментами, способный расщеплять белки, жиры и углеводы различной сложности.
Пищевой ком попадая в кишечник изначально подвергается механической обработке за счет сокращения стенок кишечника, которая носит волнообразный,маятникообразный или перистальтический характер и называется моторикой кишечника(Рис. 41-42).
В начальную часть кишечника (тонкую кишку) впадают желчный проток и проток поджелудочной железы.
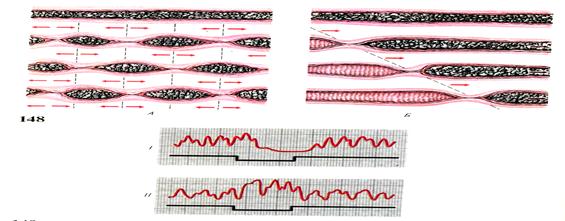
Рис.42 1) Схема сокращения кишечника рыб- моторика.
§
А — маятникообразное движение. Б- перистальтическое движение
2) Регуляция моторики кишечника; І-раздражение симпатических
По ним в кишечник попадают желчь и ферменты поджелудочной железы, под действием которых происходит расщепление белков до аминокислот, жиров до глицерина и жирных кислот и расщепление углеводов — полисахаридов до сахаров, главным образом, глюкозы.
В кишечнике помимо расщепления питательных веществ, происходит их всасывание, наиболее интенсивно протекающее в заднем участке, так называемом толстым отделом кишечника. Толстый кишечник несет преимущественно всасывательную функцию, причем в этом отделе интенсивно поглощается вода из состава химуса, а в других отделах вода, наоборот, секретируется в просвет кишечной трубки, поддерживая жидкое состояние химуса. Этому способствует складчатое строение его стенок, наличие в них ворсинкообразных выростов, пронизанных капиллярами и лимфатическими сосудами, наличие клеток, секретирующих слизь.
Сложный рельеф слизистой оболочки толстой кишки, образуемый тонкими ворсинками, пластинками, складками, увеличивает поверхность всасывания, а также происходит более детальный процесс пищеварения так называемая – пристеночное или мембранное пищеварение. В каудальной части кишечника, у многих животных морфологически выраженной в виде прямой кишки, формируются достаточно обезвоженные части химуса — фекалии, которые выделяются в клоаку, а из нее – наружу (у костистых рыб и высших млекопитающих – прямо наружу). Общая длина кишечника зависит от типа питания: сложный внутренний рельеф кишечника увеличивает поверхность всасывания.
У многих видов в начальной части кишечника помещаются слепые отростки — пилорические придатки, число которых сильно варьирует: от 3 у окуня до 400 у лососей. Карповые, сомовые, щуки и некоторые другие рыбы пилорических придатков не имеют. При помощи пилорических придатков всасывающая поверхность кишечника увеличивается в несколько раз.
У многих видов в начальной части кишечника помещаются слепые отростки — пилорические придатки, число которых сильно варьирует: от 3 у окуня до 400 у лососей. Карповые, сомовые, щуки и некоторые другие рыбы пилорических придатков не имеют. При помощи пилорических придатков всасывающая поверхность кишечника увеличивается в несколько раз.
У рыб, не имеющих желудка, кишечный тракт представляет собой большей частью недифференцированную трубку, суживающуюся к концу. У некоторых рыб, в частности у карпа, передняя часть кишечника расширена и напоминает по форме желудок. Однако это лишь внешняя аналогия: здесь нет характерных для желудка желез, вырабатывающих пепсин.
Печень – крупная, сложная трубчато-сетчатая, пищеварительная железа, по происхождению связанная с кишечником ее масса составляет у акул 14 — 25%, у костистых — 1 — 8% массы тела, по размерам уступает только гонадам. У зародышей является слепым выростом. Желчные протоки проводят желчь в желчный пузырь (только у единичных видов нет желчного пузыря). Желчь благодаря щелочной реакции нейтрализует, кислую реакцию желудочного сока. Она эмульгирует жиры, активирует липазу — фермент поджелудочной железы. Из пищеварительного тракта вся кровь медленно протекает через печень. В печеночных клетках кроме образования желчи происходит обезвреживание попавших с пищей чужеродных белков и ядов, откладывается гликоген, а у акул и тресковых (треска, налим и др.) — жир и витамины. Пройдя через печень, кровь по печеночной вене направляется к сердцу. Барьерная функция печени (очищение крови от вредных веществ) обусловливает ее важнейшую роль не только в пищеварении, но и кровообращении.
Поджелудочная железа — сложная альвеолярная железа, также производная кишечника, является компактным органом только у акул и немногих других рыб. У большинства рыб она визуально не обнаруживается, так как диффузно внедрена в ткань печени (большей частью), и поэтому ее можно различить только на гистологических препаратах. Каждая долька связана с артерией, веной, нервным окончанием и протоком, выводящим секрет к желчному пузырю. Обе железы носят общее название hepatopancreas.
В поджелудочной железе вырабатываются пищеварительные ферменты, действующие на белки, жиры и углеводы (трипсин, эрепсин, энтерококиназа, липаза, амилаза, мальтаза), которые выводятся в кишечник. У костистых рыб (впервые среди позвоночных) встречаются в паренхиме поджелудочной железы островки Лангерганса, в которых многочисленны клетки, синтезирующие гормон инсулин, выделяемый прямо в кровь и регулирующий углеводный обмен. Таким образом, поджелудочная железа является железой смешанной функции, т.е. внешней и внутренней секреции.
Из мешкообразного впячивания спинной части начала кишечника образуется у рыб плавательный пузырь — орган, свойственный только рыбам.
5.7 Внутриклеточное и внеклеточное — мембранное пищеварение.Самая простая пищеварительная система – это внутриклеточная, за счет пищеварительных вакуоли.
Внутриклеточное переваривание обеспечивается в стенках пищеварительного тракта подвижными амебоцитами, которые обладают свойствами фагоцитов и содержат определенное количество пищеварительных ферментов.
Современные представления о внутриклеточном переваривании пищи основаны на предположении об участии в этом процессе лизосом. Лизосомы – это пузыревидные органеллы цитоплазмы, содержащие набор различных гидролаз (фосфатаз, пептидаз, гликозидаз и др.).
Переваривание белков. Жиры и многие углеводы, обладающие крупной и сложной молекулой, не могут непосредственно проникнуть в кровеносные капилляры. Эти вещества предварительно расщепляются на отдельные составные части, которые и попадают в кровяное русло.
Пищеварительные эндопептидазы расщепляют как внутренние пептидные связи, так и связи, расположенные на конце молекулы белков и пептидов, а экзопептидазы последовательно отщепляют аминокислоты с аминного или карбоксильного конца.
Переваривание белков заключается в последовательном расщеплении длинной белковой молекулы до отдельных аминокислот, которые затем всасываются и из которых организм строит собственные видоспецифические белки. Ферменты, переваривающие белки, называются пентидазами, они обладают свойством разрывать пептидные связи, соединяющие молекулы отдельных аминокислот.
Имеются две группы таких ферментов, отличающиеся по месту расположения пептидной связи, на которую они воздействуют. Эндопептидазы (действуют на связи, расположенные внутри пептидной цепочки), а экзопептидазы (гидролизуют концевые связи). Эндопептидазы в свою очередь различаются по характеру гидролизуемой пептидной связи. Наиболее распространенные из них ферменты пепсин, трипсин и химотрипсин.
Пепсин специфически действует на экзопептидазы. Пептидные связи между аминогруппой ароматической аминокислоты и карбоксильной группой дикарбоновые аминокислоты.
Трипсин гидролизует пептидные связи, образованные карбоксильными группами аминокислот, содержащих две аминогруппы.
Химотрипсин разрушает связь, прилежащую к карбоксильной группе ароматической аминокислоты, т.е. его действие противоположно действию пепсина.
Пепсин вырабатывается железами, расположенными в слизистой оболочке желудка, в неактивной форме (пепсиноген). Позвоночные за исключением не имеющих желудка рыб, круглоротых и нескольких видов с нежелизистым желудком, секретируют пепсиноген, имеющий оптимум активности при рН 1,8-2,0. Сразу после поступления в желудок рН пищи сохраняется обычно выше 2 до тех пор, пока не выделится достаточное количество кислоты. В кислой среде (рН<6) пепсиноген превращается в активный пепсин. Этот фермент активен при кислой реакции среды: у млекопитающих оптимизм отмечается при рН –1 –2. Реакция среды в желудке разных животных самая разнообразная. Так, у костистых рыб значение рН желудочного содержимого колеблется от 1,8 до 6,8 (и даже до 7-8,5 у некоторых видов кефалей).
У рыб, лишенных желудка (химеры, двоякодышащие, карповые и др.), соляная кислота вообще не выделяется, отсутствует у них и пепсин. У растительноядных рыб кислотность содержимого желудка коррелирует со способностью к измельчению пищи. У видов, не измельчающих пищу, реакция среды кислая (рН 2-4), что способствует лизису клеточных оболочек. Рыбы, перетирающие пищу, характеризуются сниженной кислотностью, что достигается путем заглатывания углекислого кальция (Payne, 1978; Lobel, 1981). Наконец, не исключена динамика величины рН в желудке, связанная с ее функциональным состоянием. Например, у керчака Муохосерhalus scorpius в пустом желудке рН 4,0-7,4, а в наполненном- 2,2-2,8. Величина рН может определяться и тем, что у рыб в желудке, помимо пепсина, есть и другие ферменты.
Известно, что химотрипсиногены и трипсиногены секретируются поджелудочной железой всех позвоночных. Трипсиноген активируется энтерокиназой в кишечнике позвоночных. Сам трипсин активирует новые порции трипсиногена и все остальные протезные зимогены. Есть 2 типа специфических панкреатических белковоподобных ингибиторов трипсина: ингибитор Кунитца (основной) и ингибитор Казаля (кислый).
Трипсин выделяется в виде неактивного трипсиногена. Он переходит в активную форму в тонкой кишке под дейтвием фермента энтерокиназы, кроме того, трипсиноген, активируется в присутствии трипсина. Последнее обстоятельство приводит к быстрому наращиванию количества фермента после начала пищеварения (явление — автокатализа). Трипсин воздействует на пищу уже вошедшую в тонкий кишечник. Максимум активности этого фермента наблюдается в слабощелочной среде при рН 6,0-8,5.
Химотрипсин также продуцируется в поджелудочной железе в виде неактивного химотрипсиногена. Попадая по протокам поджелудочной железы в тонкую кишку, под действием свободного трипсина он превращается в активный химотрипсин. Оптимум активности этого фермента, сходен с трипсином. Оставшиеся части белковых молекул подвергаются действию экзопептидаз. Эти ферменты также обладают специфичностью, так карбоксипептидаза секретируемая в поджелудочной железе, избирательно отщепляет конечную аминокислоту со свободной карбоксильной группой, а аминопептидазе (секретируется в тонком кишечнике)- аминокислоту со свободной аминогруппой. Наконец, дипептидазы, также продуцируемые в тонком кишечнике, разрушают связь в ферментах, состоящих всего из двух аминокислот.
У некоторых рыб всасывание частично происходит уже в желудке, где помимо пепсина обнаружены другие протеазы. Например, эластаза выделяется с панкреатическим соком в виде проэластазы, от которой образуется фермент с молекулярным весом 26000. Эластаза расщепляет многие белки, обладает уникальной способностью переваривать эластин — фибриллярный белок. В поджелудочной железе хищных рыб, проэластаза содержится в больших количествах.
Коллагеназа, которая расщепляет неденатурированный коллаген, была обнаружена у краба. Этот фермент обеспечивает усвоение падали.
Энтерокиназа локализована в щеточной кайме кишечных ворсинок эпителиальных клеток. С помощью желчных солей фермент попадает в просвет кишечника, поэтому активация трипсиногена происходит на поверхности слизистой и в просвете кишечника.
Карбоксипептидаза В действует совместно с трипсином аналогично, а карбоксипептидаза А действует совместно с химотрипсином. Конечная стадия полного расщепления до аминокислот осуществляется аминопептидазами, три — и дипептидазами.
Большую роль в переваривании белков у рыб играют пилорические выросты — слепые отростки, расположенные между желудком и тонкой кишкой, в полости которых обнаружена высокая протеолитическая активность. По-видимому, пилорические выросты обладают также всасывательной функцией.
Переваривание жиров.Триглицериды животного и растительного происхождения представляют собой главные пищевые жиры и липазы, которые при переваривании этих веществ гидролизуют длинные цепочки эфиров, встречаются повсеместно. Панкреатическая липаза – основной фермент, расщепляющий жиры. Липаза присутствует в активной форме и секретируется у всех позвоночных. Ее субстраты эмульгируются под действием солей желчных кислот. Жирные кислоты и моноглицерины отщепляются от субстратов и всасываются эпительными клетками.
Нейтральные жиры состоят из жирных высших кислот, связанных с трехатомным спиртом-глицерином. Жиры практически нерастворимы в воде, что сильно затрудняет их ферментативный гидролиз. Этот процесс облегчается эмульгированием жиров, в качестве эмульгатора выступает желчь, выделяемая в печени. Натриевые соли желчных кислот представляют собой поверхностно-активные вещества. Обволакивая капельки жира, они снижают поверхностное натяжение на границе жира и воды и таким образом превращают жировые капли в тонкодисперсную эмульсию.
Гидролиз эмульгированных жиров начинается в двенадцатиперстной кишке под действием фермента липазы, который секретируется поджелудочной железой и активируется под влиянием желчных кислот. В результате в виде конечного продукта освобождаются глицерин и жирные кислоты.
Жирные кислоты, также почти нерастворимы в воде, поддерживаются в дисперсном состоянии с помощью солей желчных кислот. В таком виде они могут проникать сквозь слизистую кишечника. В какой-то мере жиры могут всасываться в не расщепленном, а лишь эмульгировнном виде, мельчайшие капельки жира непосредственно захватываются эпительными клетками кишечника. Глицерин, образующийся при ферментативном гидролизе жира, всасывается легко вследствие высокой растворимости. Более сложные липиды (холестерин, фофсфолипиды, воск) гидролизуются специфическими ферментами.
Расщепление углеводов.Полисахариды, входящие в пищу рыб относятся в основном к двум классам: а) структурные полисахариды, которые не перевариваются позвоночными — целлюлоза, лигнин, декстраны, маннаны, инулин, пентозаны, пектиновые и альгиновые кислоты, агар и хитин; б) универсальные пищевые полисахариды – крахмал и гликоген. Крахмал служит самым важным питательным резервным веществом растений и состоит из амилазы и амилопектина.
Углеводы представлены в пище в разных формах, и переваривание их может идти по-разному. Простые сахара (глюкоза, фруктоза) всасываются без ферментативного воздействия и прямо утилизируются. Сложные сахара как поли и дисахариды, к которым относятся сахароза (дисахарид растительного происхождения) и лактоза (содержится в молоке), перед всасыванием предварительно разлагаются до моносахаридов. Ферменты, расщепляющие дисахариды (сахароза, лактоза, мальтоза) секретируются железами тонкого отдела кишечника.
Многие животные используют в пищу части растений, богатые резервным растительным углеводом — крахмалом. Крахмал — полимер, состоящий из большого количества молекул глюкозы, в пищеварительном тракте он поэтапно расщепляется до моносахаридов.
Несмотря на очень слабую растворимость, крахмал легко гидролизуется ферментом амилазой (α- и β- формы). У большинства млекопитающих этот фермент обильно продуцируется слюнными железами, а также входит в состав секрета поджелудочной железы. У других животных как рыб, амилаза выделяется только в поджелудочной железе, хотя у некоторых птиц регистрируется слабая активность амилазы и в ротовой полости.
Гликоген — резервный полисахарид животных. Животный крахмал гликоген гидролизуется с помощью фермента гликогеназы (γ-амилаза или глюкоамилаза), обнаруживаемого в составе пищеварительных соков двенадцатиперстной кишки (реже в желудочном соке). На этом этапе образуются молекулы дисахарида — мальтозы, которая затем под действием специфического фермента расщепляется на две молекулы глюкозы. В основном же гликоген глюкозы и там же гидролизуется при снижении уровня сахара в крови.
У некоторых в желудке обнаружены ферменты хитиназа и хитобиаза, гидролизующие хитин — сложный азотсодержащий углевод. Хитиназа вырабатывается также в секрете желудочных желез и в панкреатическом соке у водных позвоночных (круглоротые и оба класса рыб), особенно у видов, корм которых содержит много хитина. Происхождение хитиназы у рыб не совсем ясно.
Конечные продукты пищеварения всасываются в разных отделах пищеварительного тракта, но наиболее активно этот процесс идет в тонкой кишке. Некоторые вещества поглощаются путем пассивной диффузии, но в большинстве случаев имеет место активный транспорт против градиента концентрации, осуществляющийся с затратой энергии.
5.8 Регуляция процесов пищеварения.Сложный характер поэтапной переработки пищи обусловливается четкой координацией моторной и секреторной деятельности пищеварительной системы, в которой участвует нервные и гуморальные механизмы.
Вервной регуляции процессов пищеварения большое значение имеет автономия висцериальной системы, представленная у позвоночных двумя нервными сетями. Так называемое ауэрбахово сплетение, расположенное между двумя слоями гладкой мускулатуры кишечника, стимулирует его спонтанную моторную деятельность. Сплетение Майсснера, локализованное в подслизистой оболочке, активирует секреторную деятельность пищеварительных желез. Автономная нервная система снабжается волокнами симпатического или парасимпатических нервов, функции которых противоположны. Баланс этих двух систем поддерживает нормальный уровень мышечного тонуса, перистальтики и секреторной активности.
У низших позвоночных механизм двойного контроля выражен неполно. В частности, у них не выявлен нервный контроль секреторной деятельности, а моторика активизируется разными нервными путями. Так, у хрящевых рыб отсутствует парасимпатическая регуляция, а стимуляция симпатических волокон возбуждает мускулатуру кишечника; у костистых рыб блуждающий нерв контролирует моторную активность желудка, а влияние симпатических волокон выражено у разных видов неодинаково. Относительно быстрое восстановление моторики желудка после ваготомии дает основание считать, что у рыб автономная регуляция выражена сильнее, чем у наземных позвоночных.
Гуморальная регуляция пищеварения в значительной мере осуществляется комплексом гормонов, секретируемых в стенках различных отделов пищеварительного тракта. Так, деятельность желудка регулирует гормон — гастрин, продуцируемый клетками слизистой желудка и стимулирующий секрецию соляной кислоты, по принципу обратной связи подавляет секрецию этого гормона.
Состав корма также влияет на уровень секреции, которая увеличивается при повышении количества белков к пище. Ряд гормонов выделяется в двенадцатиперстной кишке, все они координируются функцией разных отделов желудка и кишечника в соответствии с характером пищи и степенью ее переваренности. Так, энтерогастрин тормозит выделение желудочного сока (особенно соляной кислоты) при поступлении в кишечник жиров (что и стимулирует продукцию этого гормона). Холецистокинин, также секретируемый под действием жиров, вызывает выброс желчи из желчного пузыря. Секреция желудочного сока стимулируется блуждающим нервом, а симпатический нерв вызывает слабую секрецию слизи.
Секретин и панкреозимы регулируют деятельность поджелудочной железы: первый выделяется под влиянием соляной кислоты, попадающей из желудка, и стимулирует обильное выделение сока, бедного ферментами, но богатого солями, что снижает кислотность, второй регулирует собственно ферметативную секрецию и продуцируются под влиянием полупереваренных белков. В двенадцатиперстной кишке продуцируются гормоны (энтерокинин, дуокринин), регулирующие деятельность тонкого кишечника.
От состава корма существенно зависит и скорость пищеварения.
У пойкилотермных животных пища переваривается значительно медленнее (время прохождение ее через кишечник исчисляется днями), процесс этот в сильной степени зависит от температуры среды.
Энергетические резервы. Энергия расходуется в организме непрерывно, тогда как поступление пищи – процесс периодический. В связи с этим важно, что часть съеденной пищи не утилизируется немедленно, а откладывается в запас. У наземных позвоночных энергетические резервы запасаются в форме гликогена и жира, большое значение в качестве источника энергии у них имеют белки. Значение углеводных запасов определяется, прежде всего тем, что нервная ткань и в свою очередь головной мозг, в качестве единственного источника энергии, используют глюкозу.
Преимущество запасения гликогена заключается в том, что он может быстро мобилизоваться и расщепляться в анаэробных условиях, образуя молочную кислоту. Такое использование гликогена характерно при экстренной усиленной мускульной работе даже у высших позвоночных; у амфибий и рептилий гликогенолиз – обычная форма продукции энергии при мышечной активности. Жировые резервы мобилизуются медленнее, но жир обладает большой энергетической ценностью и широко используется в качестве источника энергии для работы различных органов, в частности мускулатуры.
Контрольные вопросы.
1. Каково назначение органов пищеварения?
2. Из каких частей состоит пищеварительный тракт рыб?
3. Каково назначение рта, его положение и форма у различных видов рыб?
4. Каково назначение и строение глотки, пищевода, желудка и кишечника рыб?
5. Пищеварительные железы рыб, их строение и назначение.
6. Основные морфологические особенности пищеварительного тракта
разных видов рыб.
7. В чем сущность пищеварительного процесса?
8. Роль механических,физико-химических,микробиологических и
ферментативных процессов в пищеварении.
9. Из каких актов слогается процесс пищеварения?
10.Какие функции выполняет желудок,назовите фазы желудочного
сокоотделения?
11.Какие функции выполняют тонкии и толстые отделы кишечника,
назовите фазы кишечного сокоотделения?
12.Какие функции выполняет печень,назовите фазы печеночного
желчеотделения, состав и функции желчи?
13.Какова роль поджелудочной железы в процессе пищеварения и механизмы нейрогуморальной регуляции?.
14.Состав желудочного и кишечного сока.
15.Каков механизм регуляции желудочного и кишечного сокращения?
ГЛАВА 6
§
ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
Основным процессом, характеризующим самую жизнь и непрерывно протекающим в живом организме, является обмен веществ. Этот процесс сводится к поступлению в организм из внешней среды различных веществ, их усвоению, изменению и выделению продуктов распада. При этом в организме осуществляется множество различных химических, механических, термических и электрических явлений. Непрерывно происходит превращение энергии, освобождение связанной химической потенциальной энергии и переход её в другие виды.
Обмен веществ отличает живой организм от мёртвого, органический мир от неорганического. Изменения веществ и превращения энергии осуществляются и в неорганическом мире.
Процессы обмена веществ состоят из явлений ассимиляции и диссимиляции, ассимиляцией называется сумма процессов созидания живой материи, т, е. усвоение веществ, поступающих в организм из внешней природы, образование сложных химических соединений из более простых. Ассимиляция — это происходящий в организме синтез живой протоплазмы.
Диссимиляцией называется изнашивание живой материи, разрушение, распад, расщепление входящих в состав протоплазмы веществ, в частности, белковых соединений.
Ассимиляция и диссимиляция протекают в организме одновременно и составляют единый процесс. (Рис. 43)
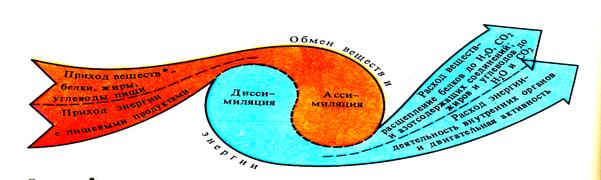
· —за исключением белков жиров,углеводов,экскрементов
Рис 43 Общее представление об обмене веществ и энергии.
Однако они не всегда взаимно уравновешиваются. В различные моменты жизни и деятельности организма один из них может преобладать.
В период роста, например, преобладают явления ассимиляции; старость характеризуется выраженным преобладанием диссимиляции.
Процессы ассимиляции и диссимиляции длительно невозможны без поступления в организм пищи и кислорода извне и выведения продуктов распада. Поэтому обмен веществ включает три этапа: 1) поступление различных органических и неорганических веществ в организм, 2) их изменение в организме и 3) выведение продуктов распада.
Источником пополнения организма веществами и энергией является пища.Пища (или корм)составляется из пищевых продуктов. Этим термином обозначается смесь пищевых (или кормовых) веществ. К пищевым продуктам (кормам) относятся, зоо- и фито — планктон. Все они содержат воду, минеральные вещества, углеводы, жиры, белки и пр. Пищевые вещества поступают в организм через пищеварительный тракт. Вместе е пищевыми органическими веществами поступает и потенциальная энергия для клеток.
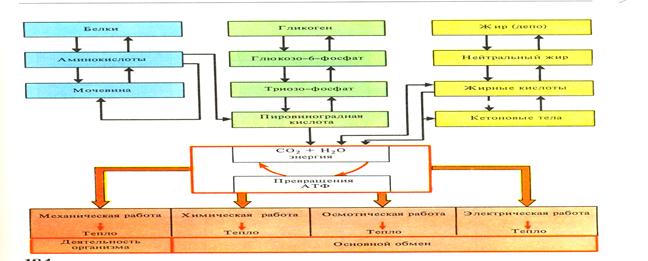
Рис.44 1) Схема превращения веществ и энергии в организме.
Характеризуя пищу с точки зрения обмена веществ и энергии, можно сказать, что пища нужна организму:
1) для построения клеток тканей организма;
2) для образования регулятора реакций — ферментов, гормонов и витаминов;
3) для пополнения организма водой;
4) как источник энергии;
5)фактор, поддерживающий на определенной высоте работу сердца, дыхательного аппарата, движение желудочно-кишечного тракта, тонус кровеносных сосудов.
Кроме того, часть пищи откладывается в организме в виде запасов (жиры, углеводы), которые необходимы организму в случае нужды, например, в период заболевания. Пища выполняет роль основной базы вещественного и энергетического обмена.
В процессе обмена веществ непрерывно происходит изменение энергии. Потенциальная (химическая) энергия органических веществ при их расщеплении освобождается и превращается в другие виды энергии в организме рыб. Обмен веществ и обмен энергии являются двумя проявлениями единого процесса. Различают обмен веществ как общий, межуточный, основной, азотистый (белковый), углеводный, жировой, минеральный и водный. (Рис. 44)
6.2 Общий обменПод общим обменом понимают, непосредственно вещественный и энергетический обмен между внешней средой и организмом при обычных условиях жизни. Этот обмен включает рассмотрение всех этапов:
1) поступление пищи и превращение её в желудочно-кишечном тракте,
2) изменение всасывающихся веществ в клетках, тканях и органах,
3) образование и выделение продуктов распада.
6.3 Межуточный обмен— это совокупность химических превращений в клетках, тканях и органах веществ, всасывающихся из кишечника.
6.4 Основной обмен— это обмен веществ в организме находящихся в состоянии полного покоя, лишённых всяких раздражений и имеющих относительно освобождённый от пищевых масс желудочно-кишечный тракт. Основной обмен, совокупность превращений в теле, которые необходимы для поддержания основных жизненных функций. Этот обмен является характеристикой интенсивности обмена веществ и энергии. Определяющими его факторами являются рост, вес, возраст и пол. Значительная часть основного обмена у теплокровных животных падает на работу дыхательной мускулатуры (10—15%), сердца (около 4%), почек (около 5%) и печени. При нормальном течении физиологических процессов величина основного обмена довольно постоянна.
В понятия общий, основной и межуточный обмены включаются процессы превращения отдельных органических веществ, воды и минеральных веществ, т.е. белковый, или азотистый, углеводный, жировой, водный и минеральный обмен.
6.5 Белковый обмен.После расщепления в желудочно-кишечном тракте белки всасываются из кишечника в кровь в виде аминокислот и несложных полипептидов. Из них в клетках синтезируется тканевой белок. По имеющимся данным, белки плазмы могут переходить в белки тканей, в частности, печени, без предварительного распада до аминокислот.
Процесс синтеза белка в организме непрерывен, потому что там постоянно происходят разрушение клеток и распад их протоплазмы. При синтезе белков необходимо известное количество различных аминокислот. Есть аминокислоты, которые не синтезируются в организме, и недостаток их в пище ведёт к нарушению белкового обмена. К таким незаменимым аминокислотам относятся триптофан, тирозин, цистин и ряд других. Биологическое значение этих аминокислот состоит в том, что при их отсутствии или недостатке наступает отрицательный баланс белка, останавливается рост, падает вес тела.
О потребности организма в белках и о размерах его распада в организме можно судить учитывая азот, введённый в организм с пищей и выведенный из организма с мочой и калом. Этим путём устанавливается так называемый баланс азота в организме.
Среднее содержание азота в белке составляет 16%. Количество введённого белка определяется путём исследования азота, содержащегося в принятой пище. Часть воспринятого с пищей азота теряется с калом, поэтому для определения усвоенного азота вычитают количество азота в кале из количества азота в пище. Найденную величину усвоенного азота умножают на коэфициент пропорциональности — 6,25, соответственно 16% содержания азота в белке (100 : 16=6,25). Таким путём узнают величину усвоенного белка в граммах. Для определения количества разрушенного белка определяют содержание азота в моче. Если азота поступает больше, чем расходуется, говорят о положительном балансе, т. е. о преобладании в организме синтеза белка над его распадом. Отрицательный баланс указывает на преобладание распада белка над его синтезом.
Задержка азота в организме, иначе ретенция его, наблюдается в период роста. С окончанием pocтa размер синтетических процессов и задержка азота резко сокращаются.
В печени может накапливаться известное количество белка, как запасный фонд, который используется организмом при недостаточном поступлении белка извне. Количество запасного белка в печени невелико. По данным исследований, значительная доля аминокислот крови поступает в тканевые элементы и также составляет там запасный фонд, особенно если эти кислоты адсорбированы в малоактивных тканях, как, например, в красных кровяных клетках, в соединительной ткани.
Одновременно с синтезом тканевого белка идёт его распад. Этот распад происходит постоянно, даже и при отсутствии поступления белка в организм. Объясняется это тем, что в результате жизнедеятельности организма всегда происходит распад некоторого количества веществ протоплазмы и разрушение некоторых клеток. Если выключить из рациона белок, а другие вещества вводить в количестве, достаточном для удовлетворения суточной потребности организма в энергии, то все же организм выделяет часть того азота, который связан с самими процессами жизнедеятельности. Эти наименьшие для организма потери белка назвали коэфициентом изнашивания.
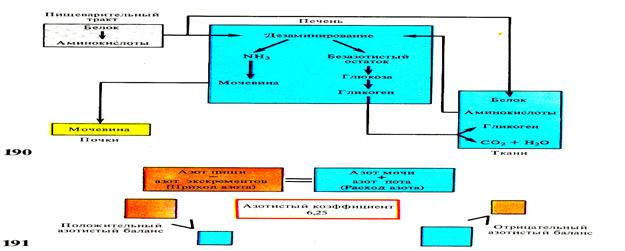
Рис.45 Схема белкового обмена, азотистый баланс
Распад белка в организме возрастает при включении его в рацион. Объясняется это, прежде всего, тем, что часть белков расходуется как энергетический материал. Освобождённый при этом азот не задерживается в организме и выделяется.
У взрослого организма количество введённого с пищей азота обычно бывает равным количеству выведенного азота. Такое состояние называется азотистым равновесием, которое служит показателем нормального белкового обмена. Организм обнаруживает сравнительно малую способность к отложению запасов белковых соединений. Поэтому при увеличенном введении белка вскоре устанавливается азотистое равновесие.(Рис. 45)
Часть белков используется на синтез в связи с распадом тканевых белков, остальное же количество служит энергетическим материалом. В результате количество усвоенных и разрушенных азотистых соединений становится равным.
Внутриклеточный распад белковой молекулы образует конечные продукты: аммиак, мочевину, мочевую кислоту и ряд других веществ, которые и выделяются с мочой. В образовании конечных продуктов белкового обмена большое значение принадлежит печени.
6.6 Обмен жиров Жиры всасываются в виде глицерина и жирных кислот. В клетках кишечника эти вещества вновь синтезируются в жировую молекулу и, чаще в виде жира данного вида животного, поступают в лимфу, а частично в кровь.
Жиры, прежде всего входят в состав клеток, как их структурная часть, в виде включений в протоплазму, и принимают участие в образовании протоплазмы, ядерного вещества и оболочки. Большое количество всосавшегося жира откладывается прозапас в жировой ткани (подкожная клетчатка, вокруг некоторых внутренних органов и внутри органов, например, в печени и мышцах).
Количество жира, входящего в структуру клеток в организме, сравнительно устойчиво, а количество запасного жира колеблется в зависимости от питания, пола, возраста, породы и конституциональных особенностей.
Жиры могут образовываться в процессе межуточного обмена из углеводов и белков. В межуточном обмене жиров большая роль принадлежит печени. Установлено, что жиры, поступающие из кишечника в кровь, задерживаются в печени, а поступающие в лимфу идут в жировую ткань подкожной клетчатки, которая составляет так называемое жировое депо организма. При усилении энергетических затрат и при голодании используется жир из депо. Существует связь между углеводами и жировым обменом в печени. При уменьшении гликогена в печени содержание в печёночных клетках жира увеличивается, и наоборот.
В процессе обмена жиры расщепляются на глицерин и жирные кислоты, которые затем подвергаются окислению. Конечными продуктами распада жира являются СО2 и Н2О.
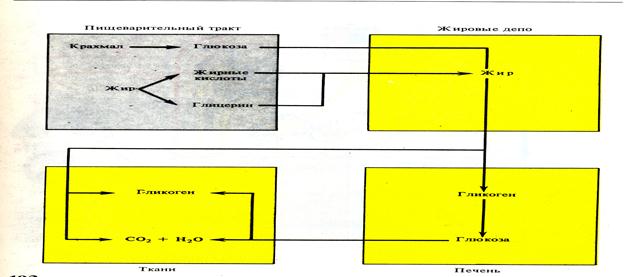
Рис.46.Схема жирового обмена (по А.Гинецинскому,1956г)
6.7 Обмен углеводовУглеводы всасываются из кишечника в кровь в виде моносахаридов, чаще в виде глюкозы (виноградного сахара). Виноградный сахар в печени синтезируется в гликоген, который составляет запасный фонд углеводов в организме. Кроме того, запасы углеводов в виде гликогена содержатся в мышцах.
Содержание сахара в крови поддерживается на определённом уровне: 0,11—0,15%. При повышении концентрации сахара выше 0,15—0,18% избыток его выделяется через почки с мочой — г л ю к о з у р и я. В нормальных условиях обмен углеводов и, в частности, содержание сахара в печени и в крови регулируются железами внутренней секреции и нервной системой. Образование углеводов может происходить в организме из белков и жиров. По мере убывания сахара в крови происходит мобилизация гликогена из мышц, благодаря чему сохраняется относительное постоянство содержания глюкозы в крови. Особенно резко увеличивается распад гликогена при мышечной работе. (Рис. 47).
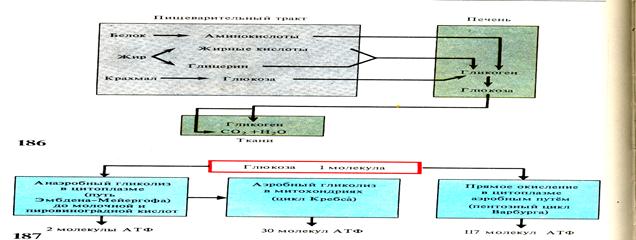
Рис.47 Схема углеводного обмена, расщепление глюкозы и образование АТФ.
Внутриклеточный распад углеводов сложен: он осуществляется сначала путём безкислородного расщепления — гликолиза, а затем путём окисления продуктов распада (молочной кислоты) до С02 и Н2О.
6.8 Обмен минеральных веществ и воды Наряду с белками, жирами и углеводами, для организма важно иметь минеральные вещества и воду. Значение их разнообразно, при их участии в организме протекают важнейшие физико-химические процессы. Благодаря минеральным солям, растворимым в воде, создаётся определённое осмотическое давление крови и тканей, сохранение которого на постоянном уровне является необходимым условием нормальной жизнедеятельности.
Минеральные вещества принимают участие в поддержании кислотно-щелочного равновесия и сохранении относительного постоянства активной реакции крови.
Вода и минеральные вещества способствуют сохранению коллоидального состояния живой протоплазмы и участвуют в построении клеточных структур. Деятельность выделительных органов, секреция пищеварительных желез и транспорт газов крови связаны с участием минеральных веществ. Значение минеральных веществ не ограничивается только их участием в физико-химических процессах. Распадаясь на ионы, они оказывают специфическое влияние на физиологические процессы. Если лишить организм воды и минеральных веществ, это приводит к тяжёлым нарушениям и смерти. Неорганические соединения в организме высших животных и человека энергетического значения не имеют. Этим они отличаются от белков, жиров и углеводов.
В обычных условиях смешанного типа питания, организм получает достаточное количество минеральных солей. Иначе обстоит дело с питанием растущего организма, который особенно нуждается в большинстве минеральных соединений.
Для развития костной и нервной ткани молоди рыб необходимо достаточное снабжение организма кальцием и фосфором. Кроме того, у растущего организма происходит задержка, натрия, калия, магния, хлора. Имеются данные, указывющие на то, что увеличение веса тела на 1 кг связано с задержкой в организме 1 г. Na.
Из кишечника минеральные вещества всасываются в виде солей и ионов. Минеральные вещества входят в состав клеток и жидкостей тела. В случае избыточного поступления солей их запас может отложиться в различных органах, а при недостатке минеральных веществ происходит мобилизация запасов. Например, кальций и фосфор откладываются в костях, железо — в печени, натрии и хлор — в коже, калий — в мышцах.
Из организма минеральные вещества выделяются с мочой, калом. С калом выделяются ионы железа, кальция и магния. Местом их выделения служит толстый отдел кишечника.
Воды в организме содержится от 35 до 63%. Содержание воды и минеральных веществ в организме подчинено регуляции через нервную систему и гуморально (через кровь).
Водный обмен организма со средой включает два противоположных процесса: поступление воды в организм и отдачу ее им во внешнюю среду. Обитатели водной среды способны получать воду через покровы или специализированные участки тканей, проницаемые для воды (например, жабры).Значение пищи в водном балансе организма заключается не только в том, что она содержит воду в тканях пищевых объектов. Усиленное питание ведет к накоплению в организме жировых запасов: в процессе окисления жиров и других органических веществ образуются так называемая метаболическая вода. Таким образом, жир в организме животных не только энергетический запас, но и внутренний источник поступления воды в клетки ткани.
Водный обмен теснейшим образом связан с обменом солей. Определенный набор солей (локов) представляет собой необходимое условие нормальной функции организма, так как соли входят в состав клеток и тканей. Играя важнейшую роль в обменных механизмах клеток, изменение количества, воды и солей нарушает осмотические процессы и ионое равновесие организма. В организме животных постоянно идет активное регулирование концентрации солевого состава жидкостей и тканей тела. Для организма водных животных важное значение имеет поддержание постоянства водно-солевого обмена, у которых осмотические процессы осуществляются не только на суборганизменном уровне, но и в зависимости от состояния окружающей водной среды.
Водно-солевой обмен у водных животных. У первично водных животных жаберные, кишечные, покровные эпителии всегда контактирует с водой, поэтому через их покровы постоянно осмотическим путем проникает вода, в том случае если концентрация солей в организме отличается от окружающей водной среды. Состав жидкостей тела рыб качественно сходен с составом морской воды. Однако количественное содержание различных ионов в теле даже морских животных может отличаться от соотношения их в окружающей их среде. Некоторые соли идут на построение скелета, а часть циркулирует в состав крови и лимфы, поддерживая постоянство внутренней среды, или, напротив, активно выводится из организма.
Ионную регуляцию в организме, связанную с поддерживанием состава и количественного соотношения различных ионов в жидкостях тела, и регуляцию осмотического давления, называют водно — солевым обменом. Изменение осмотического давления внутри организма животных приводит к изменениям давления внешней среды, таких животных называют пойкилоосмотичными (осмоконформерами). Животные, способные к активной регуляции осмотического давления поддерживают относительное постоянство внутренней среды независимо от состояния внутренней и окружающей среды, таких животных называют гомойоосмотическими (осморегуляторами).
Осморегуляция (от осмос и лат. regulo — направляю), совокупность физико-химических и физиологических процессов, обеспечивающих относительное постоянство осмотического давления (ОД) жидкостей внутренней среды (межклеточных жидкостей, лимфы, крови) у гомойосмотических животных. Механизмы осморегуляции обнаруживаются у организмов, обитающих в средах с различной концентрацией осмотически активных веществ, главным образом солей. Осморегуляция присуща всем пресноводным и наземным животным; среди морских организмов осморегуляцией обладают все позвоночные (кроме миксин) и некоторые ракообразные.
В основе физиологических механизмов осморегуляции лежит соответствующий рефлекс: изменение осмотического давления крови или межклеточной жидкости, которые воспринимаются специальными нервными клетками — осморецепторами. Возникающие в них импульсы передаются в нервные центры головного мозга, которые регулируют потребление воды и солей и их выделение осморегулирующими органами — нефридиями, почками, солевыми железами и др.
Механизм осморегуляции может быть гиперосмотическим, когда осмотическое давление внутренней среды организма больше, чем в жидкости внешней среды, и гипоосмотическим, когда внутреннее осмотическое давление меньше внешнего. В первом случае избыток воды выделяется животным главным образом через почки (растением — преимущественно через устьица); во втором — пассивную потерю воды через покровы тела организм восполняет, потребляя богатую солями воду и выделяя избыток солей в основном через солевые железы.
Пресноводные животные и морские хрящевые рыбы характеризуются гиперосмотической регуляцией. У акул и скатов это обусловлено высокой концентрацией в крови мочевины. В тело этих животных через проницаемые для воды участки покровов тела по осмотическому градиенту поступает вода. Избыток её у всех животных выделяется почками или их аналогами (сократительные вакуоли простейших, нефридии), а соли всасываются из пресной воды клетками жаберного аппарата или кожи (у земноводных).
Гипоосмотическая регуляция характерна для морских костистых рыб, морских пресмыкающихся и некоторых других. Эти животные теряют воду через покровы и с мочой. Для компенсации потерь они пьют морскую воду, опресняемую клетками солевых желёз и др. органами, выделяющими концентрированные растворы солей.
У млекопитающих основной орган осморегуляции — почки, способные выделять гипотоническую мочу при избытке воды и осмотически концентрированную — при её дефиците. Проходные рыбы (например, лососи) и некоторые ракообразные обладают обоими механизмами осморегуляции, могут жить как в пресной, так и в морской воде.
У пойкилосмотических животных (морские моллюски, иглокожие) осмотическое давление крови меняется параллельно с его изменением во внешней среде. Однако у этих животных имеется клеточная осморегуляция: при увеличении осмотического давления крови в клетках на такую же величину возрастает концентрация органических веществ (главным образом аминокислот), для которых клеточная мембрана слабо проницаема. В результате концентрация солей и содержание воды в клетке не меняются, осмотическое давление выравнивается за счёт накопления осмотически активных веществ. При уменьшении осмотического давления внешней среды и крови в клетках снижается концентрация органических веществ. Клеточная осморегуляция создаёт условия для ограниченной адаптации пойкилоосмотических животных к колебаниям осмотического давления во внешней среде.
Осморегуляция у пресноводных рыб. В организме пресноводных животных концентрация жидкостей внутренней среды обычно несколько ниже, чем у морских форм, но выше чем в окружающей воде. Пресноводные животные всегда гипертоничны по отношению к среде.
Благодаря этому вода осмотическим путем постоянно поступает внутрь организма. Концентрация солей в пресных водоемах колеблется в пределах 0-0,5%, в среднем составляя 0,2%. У некоторых пресноводных животных различные покровные образования затрудняют проникновение воды через кожу (панцири, чешуя). Однако полная изоляция организма от осмотического поступления воды невозможна.Осморегуляция организма пресноводных рыб сводится к тому, что вода поступает в организм осмотическим путем через жабры и слизистую пищеварительного тракта,а избыток ее выводится через почки. Активного питья воды не происходит. (Рис.48.)
Таблица 15 Концентрация некоторых веществ в плазме крови водных позвоночных, ммоль/л.
| Виды | Среда обитания | Na | К | Мочевина | Осмотическая концентрация мосм/л |
| Морская вода Круглоротые Миога Mijxine Минога Petromyzon Минога Lampetra Хрящевые рыбы Cкат Raja Скат Potamotrygon Акула Sgualus Кистеперые рыбы Латимерия Latimeria Костистые рыбы Золотая рыбка Carassius Рыба-жаба Ospanus Угорь Anguilla >> >> Лосось Sabmo | море море пресные воды море пресные воды море море пресные воды море пресные воды море пресные воды море | —
| —
| — — <1 <1
|
|
Соли поступают с пищей и через жабры в процессе дыхания, а потеря их идет с мочой, экскрементами и частично через кожу. Фильтрационно-реабсорбционная функция почек, абсорбция, активность жаберного эпителия, зависит от соотношения разностей осмотических давлений воды и жидкостей организма.
Осморегуляция в море – у костных рыб. В результате морской трансгрессии многие материковые водоемы оказались связанными с океаном и осолонились. Условия водно-солевого обмена в морской и пресной воде кардинальным образом отличаются, в море организм животных оказывается гипотоничным по отношению к среде, что вызывает, постоянный отток воды из организма в окружающую среду и создает угрозу обезвоживания организма.
Приспособление к осморегуляции в таких условиях развивалось на основе пресноводного принципа строения почки, свойственного у большинства морских костных рыб. В этом случае функциональная активность клубочковой системы снижается, значительная часть клубочков вообще не участвует в фильтрации. Например, фильтрационная активность отдельных нефронов у форели в морской воде выше (3,74 нл/мин против 1,31 нл/мин в пресной воде), общая гломерулярная фильтрация за счет уменьшения числа функционирующих нефронов в морской воде ниже, чем в пресной (соответственно 20,1 и 142,6 нл/мин). У многих морских костистых рыб в соленой воде уменьшается размер клубочков, и их число. Среди морских костистых рыб есть антарктические виды, совершенно утратившие клубочки, а канальцы нефронов в таких агломерулярных почках заканчиваются слепо и обладают активной секреторной функцией. Утрата клубочков, способствует сохранению в крови гликопротеидов, играющих важную роль «биологических антифризов».
Снижение уровня почечной фильтрации не в состоянии компенсировать осмотические потери воды. Поэтому морские рыбы регулярно пьют воду, получая при этом избыточное количество солей. Угорь и морской подкаменщик поглощают за сутки до 50-200 см3 воды. Если блокировать ее поступления через кишечник, рыба обезвоживается и после потери 12-20% исходный массы погибает.
Реабсорбция ионов в почечных канальцах морских рыб снижена, зато здесь происходит обратное всасывание воды из канальцевой мочи. Соответственно морские рыбы выделяют сравнительно небольшое количество мочи, концентрация которой почти равна плазме крови. Интенсивно всасывается вода и в мочевом пузыре костистых рыб, стенки которого обладают высокой осмотической проницаемостью. Здесь абсорбируется хлористый натрий.
Избыток солей, кроме почек, выводится также через кишечник, здесь происходит интенсивное всасывание воды, а соли концентрируются и выводятся с фекалиями. Это относится главным образом к двухвалентным ионам, тогда как хлористый натрий активно абсорбируется, создавая в слизистой оболочке градиент концентрации, вследствие которого вода переносится через стенку кишечника.
Большую роль в выведении избытка солей играют жабры, одновалентные (главным образом Na и Cl—) экскретируются почти исключительно через жабры, выполняющие у рыб двойную функцию- дыхания и выделения. В жаберном эпителии есть особые крупные бокаловидные клетки. Эти «хлоридные» клетки расположены в первичных жаберных лепестках и в отличие от дыхательных клеток связаны с сосудами венозной системы. Перенос ионов через жаберный эпителий имеет характер активного транспорта и идет с затратой энергии. Стимулом экскреторной деятельности хлоридных клеток является повышение осмолярности крови.
Участие жабр в активном транспорте одновалентных ионов характерно не только для костистых рыб. Хлоридные клетки обнаружены у акуловых рыб, хрящевых и костных ганоидов, свойствен и пресноводным рыбам. Гипертоничность внешней среды обусловливает постоянные осмотические потери воды (в основном через жабры), которые восполняются путем питья. Избыток получаемых при этом солей выводится через почки и фекалиями (главным образом двухвалентные ионы), а также активно экскретируются особыми клетками жаберного эпителия (в основном Na и Cl—). Функционирование механизмов осморегуляции позволяет поддерживать осмотическое давление внутренней среды на относительно постоянном уровне, гипотическом по отношению к внешней среде.
Механизм осморегуляции у хрящевых рыб. Концентрация солей в их крови близка к таковой у костистых рыб и ниже, чем концентрация их в морской воде. Однако осмотическое давление жидкостей тела у этих рыб практически равно осмотическому давлению морской воды. В почечных канальцах хрящевых рыб активно реабсорбируется мочевина, 70-90% которой возвращается обратно в кровь, повышая ее суммарное осмотическое давление. Проницаемость жабр для мочевины понижена, и избыток мочевины выводится только через почки. Помимо мочевины, в крови хрящевых рыб накапливается значительное количество триметиламиноксида (ТМАО), обладающего высокой осмотической активностью. У костистых морских рыб содержание ТМАО в крови составляет 25-460мг %, то у акуловых оно достигает 250-1470 мг%.
На снижение солености среды хрящевые рыбы реагируют уменьшением реабсорбции мочевины и усилением выведения ее с мочой. Благодаря этим регуляторным процессам акуловые рыбы выдерживают больше колебания солености среды, появляясь даже в пресных водах.
Изоосмотичность морских пластиножаберных рыб не означает отсутствия ионной регуляции. Напротив, содержание солей в составе внутренней жидкой среды поддерживается на довольно постоянном уровне. Количество хлористого натрия в крови акул и скатов почти в два раза ниже, чем в морской воде, несмотря на постоянный приток этих ионов с пищей и благодаря диффузии через жабры. Часть избыточных солей выводится с мочой, концентрация ионов калия, кальция, магния и некоторых других в проксимальных сегментах нефронов нарастает.
Хлористый натрий, напротив, реабсорбируется в дистальных почечных сегментах нефронов и возвращается в кровь. Главную роль в экскреции хлористого натрия, видимо, играет ректальная железа – пальцеобразный вырост, соединенный протоком с клоакой. Показано, что до 75% выводимого из организма хлористого натрия экскретируется ректальной железой. Акуловые рыбы в отличие от костистых не пьют морскую воду.
Значительное количество мочевины синтезируется и задерживается в крови у двоякодышащих рыб. У осетровых рыб в морской воде также увеличивается содержание осмотически активных веществ сыворотки крови, что ведет к выравниванию осмотического давления крови и окружающей среды. При этом в отличие от акуловых рыб у ряда осетровых колеблется и содержание солей, в частности хлористого натрия. Осморегуляция у осетровых рыб происходит по промежуточному типу, между осморегуляцией хрящевых и костистых рыб.
Физиологический контроль осморегуляции.Механизмы регуляции водно – солевого обмена у рыб находятся в сложном взаимодействии. Водно – солевой обмен может лабильно изменяться в зависимости от динамики условий внешней среды и внутреннего состояния организма. Адаптивные возможности проявляются у разных видов неодинаково.
Примером широкой приспособляемости к солевому режиму водоемов служат так называемые проходные формы круглоротых (миноги) и рыб. При миграциях из моря в реки (или наоборот) механизмы осморегуляции у этих животных полярно преобразуются (смена гипо – и гипертонического состояния организма), что основано на пресноводном типе структуры почек у всех водных позвоночных.
У молоди лососевых рыб процесс подготовки к смене типа осморегуляции начинается еще в реке: увеличивается концентрация осмотических активных веществ, в плазме крови возрастает число хлоридных клеток в жабрах.
Таблица 16. Показатели осморегуляции у угря Anguilla amgnikka в реке и в море (по Строганову, 1962).
| Водоем | D to среды | Выделение мочи (млкг в сутки). | D to мочи | Dto крови |
| Река | 0,08 | 60-150 | 0,09 | 0,63 |
| Море | 1,85, | 2-4 | 0,79 | 0,82 |
Все это повышает устойчивость к возрастающей солености воды при выходе в море. У взрослых особей перестройки носят обратный характер, что позволяет им подняться вверх по течению на месте нереста. Все эти изменения стимулируются нейроэндокринными механизмами.
Активная осморегуляция обеспечивает не только приспособления принципиального характера (регуляция в пресной или морской воде), но и лабильные адаптивные реакции на изменение градиента плотности. У рыб регуляция этих процессов связана главным образом с нервно-гуморальными механизмами системы гипоталамус – гипофиз – интерреналовая ткань.
Гипоталамус представляет центральную структуру головного мозга, в которой сигналы с осморецепторов трансформируются в реакции организма. Вырабатываемые в ответ на осмотическую стимуляцию нейросекреты гипоталамуса переносятся по аксонам в заднюю долю гипофиза (нейрогипофиз), откуда в виде гормонов поступают в кровь. У рыб в нейрогипофизе продуцируются гормоны аргинин – вазотоцин (АВТ), и изотоцин (ИТ), окситоцин (ОТ) и вазопрессин (ВП). Первые три гормона в низких дозах (до 1 мг/кг) действуют в пресной воде как антидиуретики, а в морской воде их влияние на диурез не проявляется. В высоких дозах (более 10 мг/кг) гормоны повышают диурез.
Гормон вазопрессин, обнаруженный у рыб, обладает диуретическим действием, причем в пресной воде он подавляет антидиуретический эффект гормонов аргинин – вазотоцина (АВТ), изотоцина (ИТ) и окситоцина (ОТ2). Все нейрогипофизарные гормоны влияют на диурез, воздействуя на интенсивность клубочковой фильтрации.
Другая форма влияния гипоталамуса на водно-солевой обмен заключается в стимуляции эндокринной деятельности передней доли гипофиза (аденогипофиз). Проникающие в аденогипофиз нейросекреты стимулируют определенные группы клеток, продуцирующих гормоны.
Наибольшее заключение в осморегуляции имеют пролактин и адренокортикотропный гормон (АКТГ). Пролактин играет роль в пресноводной осморегуляции, секретируется большем количестве у пресноводных рыб, чем у морских. Стимуляция других клеток аденогипофиза влечет за собой увеличение секреции АКТГ, которая в свою очередь повышает активность интерреналовой ткани, клетки которой выделяют ряд кортикостероидов, принимающих непосредственное участие в регуляции водно-солевого обмена. В интерреналовой ткани костистых рыб вырабатываются не менее шести кортикостероидов, а также ряд их производных. Наиболее значительную роль в регуляции водно-солевого обмена играет гормон кортизол, в меньшей степени — альдостерон. (Рис. 49).
Кортизол восстанавливает осморегуляторные свойства и считается главным фактором приспособления рыб к осморегуляции в морской воде. У акулообразных и осетровых рыб гормональный контроль осморегуляторных процессов осуществляется аналогично.
Рис. 49. Механизм гуморальной регуляции водно – солевого баланса
6.9 Обмен веществ в целом Общий обмен веществ не является постоянным и неизменным. Он значительно изменяется в зависимости от состояния организма и внешних условий.
На обмен веществ могут оказать влияние условия среды обитания рыб и характер питания, температурный режим воды и такие факторы, как возраст, рост, вес тела и пол. Функциональное состояние отдельных систем и органов также сказывается на интенсивности обмена веществ, особенно при заболевании. Ряд внешних условий, например, температура, барометрическое давление, состав и влажность воздуха, солнечная энергия и пр., может также резко влиять на обмен веществ.
О величине процессов обмена можно судить по продуктам распада, выделяемым из организма. Учёт азота в моче позволяет выяснить количество распавшегося в организме белка. Представление о распаде жиров и углеводов можно составить по выделению из организма углерода в форме СО2, так как конечным продуктом распада этих веществ является углекислый газ. Выделение СО2 из организма происходит через жабры и полужабры. Для учета выделенного СО2 и поглощённого О2 в физиологических экспериментах животных помещают в респирацион-
ную камеру. Зная количество выделенного азота и СО и поглощённого кислорода, можно составить представление об интенсивности обмена и степени использования организмом питательных веществ.
Дыхательный коэфициент— это соотношение между объёмами выделенного С02 и поглощённого О2. Он бывает различным: при окислении углеводов он равен 1, при распаде жиров — 0,7, а при распаде белков — 0,8.
Обмен веществ при голодании.При голоде организм не получает извне питательных веществ: Поддержание основных процессов, обеспечивающих жизнь организма, происходит за счёт распада запасов питательных веществ. Первый период голодания характеризуется распадом веществ, полученных организмом с воспринятой до голодания пищей.
С первых дней голодания распад белков значительно снижается, а в последующие дни расщепление белка поддерживается на низком уровне. Энергетические затраты происходят за счёт гликогена, который защищает белок от гибели. Когда запасы гликогена потрачены, организм живёт за счёт своего жира. Этот период зависит от богатства жиром данного животного. Поэтому жирные организмы голод переносят дольше, чем худые. При исчезновении запасов жира, как источника энергии, наблюдается повышенный распад тканевого белка, который организм оберегал от распада. Этот период усиленного распада тканевого белка является предвестником голодной смерти.
Во время голодания органы уменьшаются в своей массе, но не в равной степени. Больше всего исчезает жировая ткань (до 93—97%), затем следуют мышцы и железы (40—70%). Меньше теряет в весе центральная нервная система (2—3%). Таким образом, происходит сохранение органов, более важных для жизни. Смерть наступает, по некоторым данным, после потери животным 20—50% своего веса.
Обмен при голодании даёт представление о способности организма к ограничению расходования белков, как основного структурного материала клеток, а также о компенсированном и последовательном распаде запасов углеводов и жиров. Такой обмен служит показателем минимального расходования веществ, необходимых для пополнения затрат на основные процессы, поддерживающие жизнь (работа сердца, дыхательных мышц, печени, нервной системы), и на регуляцию тепла.
БЕЛКОВЫЙ МИНИМУМ.Белок пищи постоянно идёт на восстановление тканевого белка и на образование регулятора жизненных процессов, например, гормонов. В питании белок нельзя заменить ни углеводами, ни жирами, поскольку они не содержат азота. Если количество белка в пище будет убавлено ниже необходимого минимума, организм уже не сумеет сохранить азотистое равновесие, и начнёт тратить свои тканевые белки. Баланс азота в этом случае становится отрицательным.
Белковый минимум, это минимальное количество белка, которое является необходимым для существования организма. Количество белка в пище должно быть выше минимальной потребности организма в азотистых соединениях. Организм всегда должен иметь некоторый резерв, который можно использовать в случае усиления физиологической деятельности.
§
6.10.1Факторы определяющие уровень обмена веществ в организме рыб;
Содержание кислорода в среде и интенсивность обмена. Одним из эндогенных факторов, влияющих на потребление кислорода, является мышечная активность. Согласно определению «основной обмен» – это потребление кислорода при относительном покое организма.
Таблица 17- Стандартный обмен
| Рыбы | Вес тела, г | Потребление кислорода, мл/(г*ч) | Температура окружающей среды, 0С |
| Илистый прыгун | 10-15 | 0,084(в морск. воде) 0,065(в пресн. воде) | |
| Серебряный карась | 0,042 | 200С | |
| Голец | 0,109 | 200С | |
| Чукучан Карликовый сомик Карп Солик Ictalurus | 43-127 | 0,109 0,092 0,067 0,07 | 150С 200С 200С 200С |
Таблица 18 — Потребление кислорода молодью серебряного карася в период активности (I) и в условиях стандартного обмена (II) при различных температурах.
| Вид рыбы | Потребление кислорода, мл/(г*ч) | ||
| Во время полета | В покое | При оцепенении | |
| Calyptc Selasphorus | 42,4 | 0,17 |
У серебряного крася эффективность использования кислорода Е, возрастает с увеличением скорости плавания V1,65 согласно уравнению;
Е=0,031 V1,65
На интенсивность обмена влияет не только мышечная, но и другая активность, например функция половой системы в период размножения. У рыб при возбуждении потребление О2 может значительно возрастать даже без заметного усиления мышечной активности.
У высокоактивных рыб, таких, как этеостома, интенсивность обмена в несколько раз больше, чем у малоподвижных видов. Это различие обнаруживается и на тканевом уровне: у активных рыб – потребление кислорода тканью головного мозга in vitro составляет от 11 до 14 мл/(кг.мин), у менее активных морского окуня и крокера от 7,7 до 9,7 и, наконец, у совсем малоподвижных – камбалы и опсануса –от 5,6 до 6,9 мл/(кг.мин.).
Температура и время года.У большинства пойкилотермных организмов интенсивность обмена растет или снижается вместе с изменением температуры тела, причем в физиологическом диапазоне температур составляет около 2,50С.
Интенсивность стандартного обмена у рыб постепенно повышается с температурой вплоть до летального уровня. Обмен в активном состоянии может либо повышаться с температурой до определенного уровня; а затем оставаться постоянным, либо проходить через максимальное значение при температуре, выше которой рыба уже не способна совершать интенсивную мышечную работу.
Разность между уровнями обмена в покое и в активном состоянии называется физиологическим резервом активности, которая возрастает с повышением температуры до некоторого максимума, а затем падает. Как резерв, так и сама активность достигают максимума приблизительно при одной и той же температуре. Водные пойкилотермные организмы, активные в различных диапозонах температур, обладают способностью к акклиматизации. Такая адаптация позволяет животному компенсировать сезонные изменения температуры или влияние географической широты.
Кумжа наиболее активна при 150С, кара – при 250С; для сомиков такой максимум не установлен. У высокоактивных костистых рыб хорошо развит механизм температурной компенсации.
Соленость. Пресноводные рыбы обычно потребляют больше кислорода, чем морские. Слегка солоноватой воде эвригалинные организмы (например, Myfilus edulis) потребляют больше кислорода, чем в морской воде, а стеностеногалинные (например, Asferias) меньше. Потребление кислорода жабрами в малосоленой воде возрастает.
Фотопериод и физиологические ритмы. У многих животных потребление кислорода подчинено суточному или лунному ритму. Фотопериод также может влиять на обмен. При 8-часовом фотопериоде Pachygrapsus потребляет на 55% больше кислород, чем при 16 часовом. При 9 часовом периоде у ушастого окуня выше, чем при 15 часовом, если температура воды не ниже 100С.
Гормоны.Обмен веществ очень часто зависит от пола животного, но эта зависимость не всегда одинакова. Влияние физических факторов на метаболизм во многих случаях опосредуется через гормоны. Экстракты щитовидной железы усиливают обмен у амфибии. Стероиды половых желез повышают интенсивность обмена у рыб.
Развитие. Стадия развития или жизненного цикла может сама по себе влиять на обмен, независимо от размеров тела. Как правило, с возрастом уровень обмена понижается, но на эмбриональной стадии интенсивность дыхания может быть невысокой.
Размеры тела и энергетические затраты.Потребление кислорода иногда выражают в расчете на одну особь, а чаще всего пересчитывают на 1 г. или 1 кг. веса тела (влажный или сухой вес), реже на единицу количества азота. Принято также выражать интенсивность дыхания в виде показательной функции от веса тела, что означает число миллилитров О2 (при стандартных давлении и температуре), потребляемых на 1 г сухого веса за 1 ч, а Vo2— количество О2 на 1 г сырого веса (или единицу объема ткани). Общее потребление кислорода растет с увеличением размеров животного, но при расчете на единицу веса оно снижается.
Таблица 19 — Зависимость интенсивность обмена от размеров тела.
| Животные (рыбы) | Размеры тела | Потребление О2 мл/(г*ч) |
| Лосось Oncorhuncus | 3-1400 | 0,97 (в акт.атм.) 0,78 (станд.обмен) |
| Серебряный карась | 100-400 | 0,85 |
| Серебряный карась | — | 0,86 |
| Cadus | — | 0,70 |
У некоторых животных кривая, отражающая зависимость интенсивности обмена от содержания кислорода в среде, становится более пологой при величинах Ро2 выше атмосферного уровня (т.е. выше 155 мм.рт.ст.). Эти животные занимают промежуточное положение между формами с зависимым и независимым типами дыхания. Характер зависимости обмена от Ро2 и положение критической точки Ркрит.. определяются рядом внешних и внутренних факторов, в том числе экологическими особенностями животного. При значениях Ро2 ниже Ркрит.
интенсивность обмена зависит от количества доступного кислорода; если даже Ро2 выше Ркрит., то лимитирующим фактором может быть концентрация субстратов, ферментов или кофакторов. Величины Ркрит. не во всех случаях постоянны, они могут несколько раз изменяться в зависимости от условий определения.
Зависимость обмена от давления О2, и механизмы этого влияния. Во время активности уровень обмена в большей степени зависит от Ро2, чем в период покоя. Максимальная скорость плавания, так же как и потребление кислорода, коррелирует с содержанием О2 в воде. У ушастого окуня величина Ркрит. растет с температурой, и при резком падении Ро2 она бывает выше, чем при медленном понижении. У рыбы Callionimus дыхание зависит от парциального давления О2: при снижении внешнего Ро2 частота дыхания и сокращении сердца уменьшается и потребление О2 падает. Рецепторы, чувствительные к кислороду, находятся в области жабр.
6.11 Теплообмен и терморегуляция Теплота — основа химических реакции, из которых складывается жизнедеятельность организма. Температурные условия оказываются одним из важнейших факторов, влияющих на интенсивность обменных процессов. Закономерность воздействия температуры на метаболизм выражается общим для всех химических реакций правилом Вант-Гоффа, согласно которому повышение температуры ведет к пропорциональному возрастанию скорости реакции.
В живом организме химические процессы всегда идут с участием сложных ферментных систем, в результате чего увеличивается их скорость и количественно меняется реакция на повышение температуры. Верхний температурный порог жизни теоретически определяется температурой свертывания белков. Необратимые нарушения структуры белков возникают при температуре порядка 600С, которую принято называть температурой «тепловой смерти».
Аналогичные нарушения метаболических и регуляторных процессов наступают и при очень низких температурах, нередко определяя собой нижний температурный предел жизни. Нарушения деятельности сердца при слабом охлаждении проявляются в ритме сокращений и сократимости сердечной мышцы, а при более сильном – в ее проводимости и возбудимости. При одном и том же снижении температуры удлинение периода диастолы выражено сильнее, чем систолы.
В результате различной реакции на температуру по достижении определенного порога (неодинакового у разных видов) возникает дискоординация функции в организме, ведущая к его гибели. При низкой температуре появляются структурные изменения в клетках и тканях, замерзание внеклеточной и внутриклеточной жидкости. По принципиальным особенностям теплообмена различают две крупные экологические группы животных: пойкилотермные и гомойотермные. К пойкилотермным (от греческого «poikilos» – «изменчивый», «меняющийся») относятся все животные, кроме птиц и млекопитающих.
Одно из наиболее заметных свойств пойкилотермных животных – неустойчивость температуры их тела, меняющийся в довольно широких пределах в зависимости от изменений температуры окружающей среды.
Температура тела. Характерная особенность теплообмена пойкилотермных животных заключается в том, что благодаря относительно низкому уровню метаболизма. Главным источником поступления тепловой энергии у них является внешнее тепло. Этим и объясняется зависимость температур тела пойкилотермных от температуры окружающей среды. Изменчивость температуры тела влечет за собой изменения скорости обменных реакции пойкилотермных животных.
Противоточный теплообмен. Теплообмен водных, наземных представлен противоточным теплообменом. В этом случае сосуды, по которым проходит холодная венозная кровь и возвращающаяся из плавников к телу, проходят рядом с артериями. Благодаря такому расположению артериальная кровь отдает тепло венозной крови, которая нагревается, прежде чем распределяться по телу. Кровь по артериям и венам течет в противоположных направлениях и между этими параллельными системами трубок происходит обмен тепла, получивший название противоточного теплообмена.
Подобная противоточная система существует между воздухом и кровью в жаберных лепестках рыб, благодаря чему артериальная кровь насыщается кислородом, сколько имеется ее в протекающей воде. Вода и кровь текут в противоположных направлениях. В результате этого рыбы извлекают из воды 80-90% содержащего в ней кислорода и создают условия для большего насыщения крови кислородом.
Энергетические затраты. Процесс метаболизма возрастает при активной мышечной работе у рыб в момент движения растраты 0,2 на 1 гр (кг) веса составит около 78 мл/км.
Энергетические затраты при плавании очень низкие вследствие того, что удельный вес тела близок к удельному весу воды, а энергия затрачивается только на преодоление сопротивления воды. При повышении жидкости в организме повышается сопротивляемость воды, что приводит к увеличению затрат энергии. У мелких рыб затраты энергии больше чем у крупных. Благодаря своей форме и площади тела, крупные рыбы легко передвигаются с меньшими усилиями.
Рис. 50 Схема регуляции температуры тела у теплокровных животных ( по Кулланде,1970).
В момент активного плавания в мышцах тунцов и акул создается высокая температура, хотя температура тела у рыб очень близка к температуре окружающей воды. Кровь, через жабры обогащаясь кислородом, охлаждается до температуры воды, вследствие того, что артериальная кровь от жабр течет по сосудам лежащим близко к поверхности тела рыбы. В толщах мышц, где температура тела, может быть на 140С выше температуры воды, находятся сосуды, поддерживающие внутреннюю температуру.
В процессе обмена веществ непрерывно происходит изменение энергии. Энергия сложных химических соединений (потенциальная) при расщеплении освобождается и превращается в тепловую энергию. Часть энергии в организме превращается в механическую и незначительное количество в электрическую.
Контрольные вопросы.
1.Дайте определенике понятий «аноболизм» и «катоболизм», их этапы,
единство и различия.
2.Охарактеризуйте роль желудочно-кишечного тракта,печени, почек и мышц
в обмене веществ.
3.Что такое «азотистый баланс»?
4.Перечислите особенности и основные этапы углеводного, жирового,
белкового, солевого, минерального, водного обмена у рыб.
5.Охарактеризуйте механизм превращения энергии в организме рыб
6.Что такое прямая и непрямая колориметрия?
7.Что такое основной обмен, как он определяется?
8.Какие факторы влияют на интенсивность теплопродукции и теплоотдачи?
9.Механизм регуляции обменных процессов.
ГЛАВА 7
§
Наличие биологических групп рыб внутри популяции обусловлено условиями ареала. Внутрипопуляционная дифференциация рыб не проявляется при отсутствии необходимых условий для ее развития. Таким образом, дифференциация в пределах популяции – это приспособление рыб к условиям размножения и развития. Внутрипопуляционная дифференциация – это важнейшая видовая адаптация к условиям ареала, что приводит к наиболее полному использованию его возможностей.
Сохранение внутрипопуляционной дифференциации у рыб, особенно важно в современных изменившихся условиях их обитания в естественных водоемах под влиянием отрицательных природных и антропогенных факторов, наблюдаются ухудшения их физиологического состояния.
7.1.Морфология и физиология системы органов выделения. Система органов выделения, основу которой у всех рыб составляют почки, не только выводит продукты распада азотистых веществ, но и обеспечивает физико-химическую устойчивость внутренней среды организма: осмотического давления, кислотно-щелочного ионного равновесия. В этих процессах участвуют почки, жаберный аппарат, кожа, пищеварительная трубка и печень. Эти системы различаются у хрящевых и костных рыб.
Выделительная система рыб представлена двумя почками, расположенными по бокам позвоночника на спинной стороне полости тела. Вдоль почек тянутся мочеточники, которые на заднем конце сливаются и один проток, открывающийся в мочевое отверстие на мочеполовом сосочке.
Рис.51. Схема морфологического формирования выделительной
§
Почки рыб – специализированный орган для выделения воды, продуктов обмена, солей и других веществ из крови организма в окружающую среду. Они располагаются под позвоночником и над плавательным пузырем, распространяясь почти до анальной области. В ходе эволюционного развития рыб наблюдаются два основных механизма мочевыделения, секреторный и фильтрационный.
Функцию почек в процессе мочеобразования общем виде можно описать так, осмотически поглощенная вода и вода метаболических процессов попадает в кровяное русло. Из спинной аорты кровь течет по сегментальной артерии под давлением в клубочек, проходя в сосудистом сплетении клубочка, теряет часть воды и некоторые растворенные в ней вещества, здесь происходит фильтрование. Этот безбелковый ультрафильтрат, или первичная моча, поступает в мочевой извилистый каналец, где происходит абсорбция нужных организму веществ – сахаров, РО4 и Cl обратнов кровь. На выходе из канальца жидкость уже обеднена растворенными веществами (конечная моча), она поступает в общий выводной проток (мочеточник).
Пресноводные рыбы находятся в гипотонической среде и поэтому вода постоянным потоком проникает в их тело через жабры, кожу и пищей. Одновременно соли выводятся с мочой, фекалиями и через кожу.
Моча, выделяемая рыбами, содержит соли и органические вещества. Избыточная вода удаляется почками, а соли реабсорбируются в почечных канальцах. Количество выделяемой мочи служит показателем интенсивности работы пресноводной почки. Количество мочи у пресноводных рыб значительно больше, чем у морских. Пресноводные рыбы должны постоянно выделять воду, чтобы не разбухнуть. Воду они могут выделять только с мочой.
Выделительная система хрящевых и костистых рыб в зависимости от места обитания имеют свои морфо-физиологические особенности, так у хрящевых рыб (пластикожаберные), представители акулы и различные скаты обитают в основном в море, лишь немногие виды найдены в пресной воде. Содержание солей в плазме морских пластикожаберных почти равно их содержанию в плазме костных рыб.
Осмотическое давление создается в организме рыб главным образом за счет накоплений большого количества мочевины и некоторого количества триметрамиколсида во всех тканях и жидкостях тела. Это позволяет акуловым рыбам поддерживать в морской воде такой же осмотический градиент, что и в пресной, выделять мочу, гипоосмотическую по отношению к крови.
Почки (ren) акулы — парные, сильно вытянутые, лежат по бокам позвоночника примерно от уровня грудных плавников до клоаки. Почки акуловых рыб относятся к мезонифрическому типу. В заднем отделе почек на их брюшной поверхности имеются тоненькие трубочки – мочеточники.
Они открываются в полость вдающегося в клоаку мочевого или мочеполового сосочка, на вершине которого имеются отверстия.
Как и у многих морских костных рыб, у акуловых почки представляют собой главный путь выделения магния, фосфатов и сульфатов, моча гипоосмотична по отношению к крови, NaCl выводится внепочечным путем.
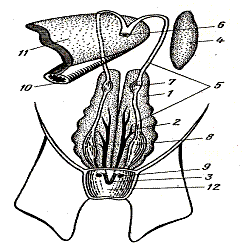
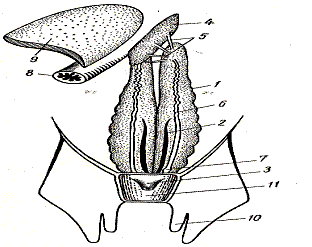
§
Почка,2- мочеточник, 3-мочеполовой сосочек, 4-левый яйчник (семеник), 5- яйцевод (семявыносящие канальцы), 6- воронка обеих яйцеводов (семяпровод), 7-скорлуповая железа (семенной пузырек), 8- матка 10-пищевод, рис. В), 9- отверстие яйцевода (печень), 10-рис. В копулятивный орган, 11-12-полость клоаки.
А В
Рис. 52. Схема мочеполовой системы А- самки, В-самца акулы:
Выведение солей NaCl из организма осуществляется ректальными железами, веделяющими секрет. Секрет данной железы у акулы, изоосмотичен по отношению к крови, но содержит более значительное количество NaCl и не содержит практически Ca2 , Mg2 и мочевины.
У костных рыб выделительная система представлена почками (ren) располагающимися на спинной стороне полости тела по обе стороны от позвоночника. По их краю проходят мочеточники (ureter), которые, выйдя из почек, сливаются в единый непарный мочевой проток. Мочевой пузырь (vesica urinaria) представляет собой вырост передней стенки начальной части этого протока. Непарный мочевой проток открывается наружу мочевым отверстием позади полового отверстия.
Парные мезонефрические (туловищные) почки костных рыб лежат ниже позвоночного столба, почти вдоль всей полости тела. У многих видов в задней половине правая и левая почки сливаются вместе. Вольфовы каналы выполняют функцию мочеточников. По выходе из почки мочеточники правой и левой почек сливаются, образовав тонкостенный полый вырост — мочевой пузырь (vesica urinaria), открывающегося наружу мочевым отверстием (у двоякодышащих — в клоаку). Почки получают артериальную кровь из почечных артерий; большие порции венозной крови приходят сюда по воротным венам почек из хвостового отдела. У пресноводных рыб хорошо развиты боуменовы капсулы с крупными клубочками. У морских костных рыб размеры клубочков уменьшаются, а у нескольких десятков видов из разных отрядов (жабы-рыбы, некоторые морские иглы и др.) клубочки практически редуцированы, такие почки называются — агломерулярными почками.
У костных рыб осмотическая концентрация крови равна 130-170 моль/см; моча у них сильно разбавлена и образуется в большом количестве. Ниже приведены данные для щуки:
| Среда, ∆нар | Кровь, ∆ВН | Моча, ∆М | Объем мочи, мл/(кг*сут) |
| -0,03 | -0,57 | -0,08 | 200-400 |
Пресноводные рыбы поглощают очень мало воды. Кожа у них относительно непроницаема для воды, однако вода осмотическим путем может проникать через жабры и слизистую рта.
Поступающая в организм вода создает в тканях рыб определенную водную нагрузку; почки пресноводных рыб имеют хорошо развитые гломерулы, которые могут фильтровать значительные объемы жидкости. По мере проведения фильтрата по канальцам большинство растворимых в нем веществ реабсорбируются, в результате чего моча становится менее концентрированной. Концентрация этой разведенной мочи, выше концентрации пресноводной среды; некоторая потеря солей происходит также за счет диффузии и в малых количествах с фекалиями. Потеря солей частично восстанавливаются поступлением их с пищей; поглощаются из среды клетками жабр.
В океане костные рыбы сталкиваются с необходимостью сберегать воду и выделять соли, поэтому выделяющие воду гломерулярные почки им не нужны. Концентрация крови у морских рыб выше, чем у пресноводных (∆ви=-0,70С). Их моча гипоосмотична по отношению к крови и образуется в очень небольших количествах.
Морские рыбы в отличий от пресноводных заглатывают некоторое количество воды. По мере того как вода проходит по кишечнику из нее поглощаются вода и некоторые соли, Na и Cl—, Mg2 и SO42-. Морские костные рыбы выделяют немного меньше мочи, так как имеют менее развитые почки, так, в почках взрослых особей опсакуса и морского черта отсутствуют гломерулы. В типичных почках морских костных рыб отсутствуют также дистальные извитые сегменты канальцев. Скорость фильтрации довольно низка даже у млекопитающих. Выделение мочи в процентах веса тела в сутки у угря равно 1,5; у камбалы –1,4.
Некоторые рыбы с осмогломерулярными почками могут адаптироваться к обитанию в пресной воде, при этом объем мочи у них увеличивается, а ее концентрация становится меньше.
Деятельность почек во многом зависит от свойств окружающей среды: находясь в морской воде, организм реабсорбирует в большей мере воду, а находясь в пресной воде, в большей мере соли. Имеются сведения и о реабсорбции некоторого количества растворенных веществ и воды в мочевом пузыре. При перемещении рыбы из морской воды в соленую функции почек изменяется. У некоторых морских рыб, которым приходится зимой переносить холода, при низких температурах происходит изменения осмотических и ионных свойств. У обитающих близко к поверхности, где образуются лед, кровь замерзает при –1,5, -1,80С. зимой и при –0,7; -0,80С. летом; понижение температуры замерзания обусловлено наличием в крови солей растворенных органических веществ. В целом в крови морских рыб при низких температурах обнаруживаются более высокие концентрации ионов, чем при высоких температурах, у пресноводных рыб концентрация ионов крови зимой или при акклимации к холоду ниже, чем летом. Костные рыбы обладают весьма богатыми возможностями осмотической регуляции.
Функция гормонов аргинин – вазотарина состоит в стимуляции обратного всасывания воды в мочевом пузыре. Механизм образования мочи заключается в следующем, у пресноводных рыб гормон аргинин-вазоторин (АВТ) повышает скорость гломерулярности фильтрации мочи. Таким образом,
в клубочке создается сила кровяного давления, которая вытесняется из капилляров в просвет боуменовой капсулы жидкую часть плазмы крови, откуда она попадает в извитой каналец нефрона. В состав этой жидкости (первичная моча) входит не только вода, но и растворенные в ней вещества, частицы которых могут проникнуть сквозь стенки капилляра (ионы солей, молекулы сахаров, мочевина, мочевая кислота), лишь белки, молекула которых слишком крупна, не попадают в состав первичной мочи. Процесс образования первичной мочи называют ультрафильтрацией. (Рис. 55-56).
В извитом канальце нефрона, который оплетен сетью кровеносных капилляров, происходит обратное всасывание солей, сахара и других веществ в кровяное русло. В результате образуется вторичная или конечная моча гипотоничная по отношению к плазме крови. (Рис.54)
В целом клубочковая система почки функционирует по фильтрационно-реабсорбционному типу, выделяя гипотическую мочу, что соответствует принципам осморегуляции в пресной воде.
Клубочковая система- главная морфологическая основа интенсивной фильтрации и выведение воды из организма. Воронки с их ресничками выводят воду со значительно меньшей скоростью.
Таблица20. Суточное количество мочи (мл/кг), выделяемое различными водными позвоночными (по Строганову, 1962).
| Виды рыб | Количество мочи, мл/кг |
| Пресноводные: Круглоротые (речная минога) Хрящевые рыбы (пила – рыба) Костные ганоиды (панцирная щука) Костистые рыбы Морские: Круглоротые (морская минога) Хрящевые рыбы (акулы) Костистые рыбы | 50-326 2-49 |
Благодаря клубочкам пресноводные рыбы выводят очень большое количество слабоконцентрированной мочи. Табл. 20
Организм непрерывно теряет часть солей в составе мочи и экскрементов, а так же в результате диффузии через кожу. Восстановление необходимых солей происходит с пищей. Важную роль в солевом обмене играет жаберный эпителий, клетки которого способны к активной абсорбции солей (ионов Na и Cl ) с окружающей воды.
Контрольные вопросы.
1.Что такое экскреция?
2.Роль почек в регуляции водно-солевого баланса.
3.Особенности строения почек у костистых и хрящевых рыб.
4.Каков механизм образования и выведения мочи?
5.Количество, состав и свойства мочи.
6.Регуляция процессов мочеобразования и мочевыведения.
7.Охарактеризуйте роль кожи в выделительных процессах.
Глава 8
8.1Общие принципы эндокринной регуляции. Эндокринные органы –органы внутренней секреции (от греч. endo –внутри, krino- выделяю), выробатывают и выделяют в кровь биологические активные вещества – гормоны, осуществляющие гуморальную регуляцию функции органов и организма в целом. К органам внутренней секреции относятся специализированные образования – эндокринные железы, а также компактные группы эндокринных клеток, расположенные в разных органах (поджелудочной железе,кишечнике,головном мозге,гонадах и др.). Эти железы небольшого размера, обильно кровоснабжены, не имеют выводных протоков для инкрета, стенки снабжены миоэпителиями способных вырабатывать секрет непосредственно в кровь.
К железам внутренней секреции относятся: гипофиз, щитовидная железа, паращитовидные железы,тельца Станниуса, интерреналовые тельца, надпочечники.
Гормоны (от греч. hormao — возбуждаю) –эта группа химических регуляторных веществ, выделяемые органами внутренней секреции в кровь, и названные Стерлингом гормонами, распространяются по всему организму, благодаря чему могут действовать одновременно на несколько органов и действие их длится более продолжительное время. Химическии активные вещества, способные в малой дозе активизировать или угнетать работу отдельных органов или систем в целом принято было называть гормонами или инкретами. К гормонам следует относить только регуляторные соединения, которые выделяются в кровь и действуют дистантно — на расстоянии и контактно. Наиболее правильно называть гормонами все химические вещества, которые вырабатываются специализированными клетками и оказывают регулирующее действие на другие клетки независимо от того, далеко расположены они или близко, как доставляются они клетками мишеням кровью или внеклеточной жидкостью путем диффузии. Одни и те же соединения могут оказывать действие как на далеко расположенные клетки через кровь, так и на близко расположенные через межклеточную жидкость.
Клетки, вырабатывающие гормоны, следует называть эндокринными регуляторными клетками, а совокупность их эндокринной регуляторной системой. Способностью к внутренней и внешней секреции, обладают только клетки эпителиальной ткани.
Железы эндокринной системы не имеют выводных протоков и секретируемые продукты-гормоны как бы впрыскивают в кровь и лимфу.Эндокринные железы встречаются двух видов: выполняющие только эндокринную функцию (щитовидная железа, надпочечники, интерреналовые тельца, гипофиз и.т.д.) и выполняющие смешанную функцию (поджелудочная железа и половые железы), которые кроме гомонов вырабатывают другие секреты.
Эндокринные клетки далеко не во всех случаях сконцентрированы в виде компактных желез, они могут быть рассеяны в разных органах даже у высших животных. На существование населенных эндокринных клеток впервые обратил внимание Фейртер. Он заметил, что некоторые рассеянные в органах эндокринные клетки регулируют деятельность близко расположенных клеток путем диффузии, и назвал их паракринными.
Делят гормоны по расстоянию от места выработки до клетки мишени на гормоны дальнего действия, переносимые кровью, и гормоны близкого действия, достигающие клетку мишень путем диффузии. Первые следует именовать телегормонами, а производящие их клетки телекринными; вторые — парагормонами, а секретирующие их клетки паракринными.
Гормоны, продуцируемые нервными клетками, естественно именовать нейрогормонами, а гормоны, вырабатываемые в кишечнике, энтерогормонами. Медиаторы по существу являются продуктами нейросекреции и отличаются от признанных нейрогормонов лишь тем, что выделяются окончаниями аксонов в синоптическую щель. Вся нервная система, и центральная и периферическая, является мощной фабрикой нейрогормонов.
Открытие гипоталамической нейросекреции сначала у рыб, а затем у высших позвоночных, разрешило гипотезу связи мозга с аденогипофизом. Деятельность всей эндокринной системы контролируется мозгом частично через вегетативные нервы, частично при посредстве либеринов и тропных гормонов аденогипофиза. Сам мозг подвергается регуляторному влиянию со стороны эндокринных органов.
Под контролем гормонов находятся все основные процессы жизнедеятельности, все этапы индивидуального развития рыб, все виды клеточного метаболизма. Они обеспечивают нормальный рост и развитие рыб, поддержание гомеостаза, размножение, адаптацию к факторам окружающей среды.
В животном организме нет двух раздельных координационных систем нервной и эндокринной, а есть лишь коммутативный регуляторный интегративный аппарат, деятельность которого осуществляется двумя, способами: проводниковым и гормональным. Ацетилхолин, выполняет функции медиатора, здесь вместе с ним должна присутствовать и холинэстераза.
У животных, вырабатывающих яд, в органах содержится гормон серотонин. Кроме серотонина из триптофана образуется мелатонин – гормон эпифиза, способствующий концентрации пигмента вокруг ядра меланиновых клеток, что приводит к побледнению или потемнению пигментных клеток рыб.
Рис.57. Схема функциональной взаимосвязи эндокринных желез
на примере млекопитающих:
§
Ядра гипоталямуса; 2-передняя доля гипофиза;3-средняя доля гипофиза;4-задняя доля гипофиза.
 8.2Щитовидная железа. Тироксин и трийодтиронин – гормоны щитовидной железы. Известное свойство тирозина – его способность присоединять йод, участвующий в регуляции обмена веществ
8.2Щитовидная железа. Тироксин и трийодтиронин – гормоны щитовидной железы. Известное свойство тирозина – его способность присоединять йод, участвующий в регуляции обмена веществ
Клетки щитовидной железы, производящие тироксин и трийодтиронин — это тирроидные фолликулы, рассеяные вдоль спинной аорты, в тканях селезенки, стенках спинной аорты. Данные фолликулы выделяют три гормона; -Т2;-Т3;-Т4. Они происходят из другого зачатка, чем те, которые вырабатывают кальцитонин. Консолидация однородных элементов в единый орган и ассоциация разнородных желез в одну, служит целям лучшей регуляции их деятельности. Паращитовидная железа появилась на уровне амфибии, у рыб она отсутствует.
Паращитовидная железа появилась только на уровне амфибии,у рыб она отсутствует
8.3Поджелудочная железа. Рассеянные вдоль кишечника эндокринные клетки занимают большую поверхность и дальнейшее разрастание их шло бы в ущерб поверхности всасывания. Поджелудочная железа, выполняет двойную функцию – секреторную и инкреторную (смешанную). Инкреторная функция заключается в выработке островками Лангерганса гормона инсулина и его антогониста глюкагона. Гормон инсулин регулирует углеводно – жировой обмен, понижая уровень сахара в крови, а глюкагон выполняет противоположную функцию.
У костистых рыб она обьединена с печенью и носит название – гепатопанкриеса, у хрящевых рыб представлен, как компактный орган. Клетки эндокринной системырыб объединяются с поджелудочной железой или образуют отдельные железки, состоящие из островковой ткани. Примером ассоциации является объединение хромаффинной и интерренальной тканей в одну железу — в надпочечник. У хрящевых рыб оба органа еще разъединены. Интерренальная железа расположена между почками, почему и получила свое название. У костистых рыб оба органа уже расположены в области кардинальных вен и соприкасаются друг с другом.
8.4Интерреналовые тельца являются аналогами корковой части надпочечников у высших животных. Морфологически они вкраплены в ткани головной почки. В интерреналовой ткани костистых рыб вырабатывается не менее шести кортикостороидов, а также ряд их производных. Наиболее важное значение в регуляции водно-солевого обмена играет кортизол и кортикостерон, в меньшей степени – альдостерон. Кортизол – выполняет стрессоподавляющую функцию, усиливает распад и накопление гликогена в печени, поддерживая постоянство концентрации глюкозы в крови. У пресноводных рыб данный гормон регулирует водно – солевой баланс, восстанавливая осморегуляторные свойства, который считается главным фактором приспособления рыб к осморегуляции в морской воде. У акулообразных и осетровых рыб гормональный контроль осморегуляторных процессов осуществляется аналогично. Кортикостерон участвует в осморегуляции, регулирует работу почек
8.5Гормоны надпочечников Альдестерон у круглоротых по – видимому, отсутствует, обнаружен лишь у некоторых рыб, в том числе у южноамериканской двоякодышащей рыбы. Деление кортикоидов на глюкокортикоиды и минералокортикоиды правомерно только для высших позвоночных. У низших позвоночных, глюкокортикоиды выполняют также и минералокортикоидную — смешанную функцию.
Определенную роль в регуляции водно-солевого обмена у рыб играют гормоны, выделяемые клетками хромаффиновой ткани, особенно адреналин (эпинефрин). Адреналин увеличивает проницаемость жаберного эпителия для воды, одновременно снижается выведение хлоридов и натрия через жабры, исполняет роль стресс гормона, выбрасываясь в кровь во время сильного стресса.
У морских рыб кортикоиды стимулируют функцию «хлоридных» клеток, удаляющих соль из организма. У пресноводных рыб, которые извлекают соль из наружной среды, они способствуют этому процессу и вместе с тем помогают задерживать натрий в организме. В почках они способствуют реабсорбции натрия.
8.6Гормоны гипофиза. Гипофиз, участок головного мозга, выполняет функцию желез внутренней секреции, является мотором всей эндокринной системы в целом. Морфологически гипофиз делится на три отдела у высших позвоночных животных, а у рыб различают два отдела, среднего отдела у рыб нет.
У позвоночных нейрогормоны выделяются преимущетсвенно в области терминалей дендритов и аксонов гипоталямо-гипофизарной системы. Поступая в кровь, спинномозговую и тканевую жидкость. Посредством этих жидкостей нейрогормон достигают клеток- мишеней и оказывают на них регулирующее влияние это аргинин – вазотоцин (АВТ), изотоцин (ИТ) и окситоцин (ОТ-2).
Все нейрогипофизарные гормоны влияют на диурез, воздействуя на интенсивность клубочковой фильтрации. Другая форма влияния гипоталамуса на водно — солевой обмен заключается в стимуляции эндокринной деятельности передней доли гипофиза (аденогипофиз). Нейросекреты, проникающие в аденогипофиз стимулируют определенные группы клеток, продуцирующих гормоны. Наибольшее значение в осморегуляции имеют пролактин и адренокортикотропный гормон (АКТГ). Адренокортикотропный гормон также регулирует процессы адаптации к стрессам, являясь антагонистом к действию адреналина. У некоторых рыб гормон пролактин вызывает утолщение эпителия и выделение кожной слизи. Кроме того, гормон поддерживает инстинкт заботы о потомстве, даже у живородящих рыб. Пролактин имеет значение для поддержания водно- солевого равновесия. Удаление гипофиза эвригалинных карпозубых рыб делает их неспособными жить в пресной воде из-за чрезмерной потери ионов натрия жабрами и через кожу, что приводит к падению осмотического давления крови. Введение пролактина спасает рыб от гибели. Пролактин играет ведущую роль в пресноводной осморегуляции. Гормон пролактин секретируется у пресноводных рыб в большем количестве, чем у морских.
Задней долей гипофиза (нейрогипофиз), вырабатывается гормон- окситоцин (ихтиотоцин), обладающим осморегуляторной и свойствами регуляции образования слизи. Вазопрессин – вазотоцин, регулирующий работу сердечно- сосудистой системы, тонус кровеносных сосудов и водно — солевой обмен. Меланофорный гормон (МФГ) — гормон регулирующий пигментацию, обычно обеспечивает темную окраску кожи.
Соматотропный гормон (СТГ) – регулирует рост и развитие организма, регенерацию тканей и органов.
Тиреотропный гормон (ТТГ) – регулирует работу щитовидной железы
Стимуляция других клеток аденогипофиза влечет за собой увеличение секреции АКТГ, в свою очередь повышающего активность интерреналовой ткани, клетки которой выделяют ряд кортикостероидов, принимающих непосредственное участие в регуляции водно – солевого обмена.
Помимо интерреналовой и хромаффиновой ткани, в почках лучеперых рыб обнаруживаются отдельные группы железистых клеток, заключенных в соединительнотканные капсулы – тельца Станниуса. Эти образования участвуют в регуляции обмена кальция и водно – солевого обмена, вырабатывают вещество гипокальцин.
Для рыб характерна еще каудальная нейросекреторная система: она находится в заднем отделе спинного мозга, где расположены нейросекреторные клетки, отростки которых оканчиваются в его брюшной части. Вырабатывает гормон ренин, регулирующий кровянное давление – ангиотензин 2.
Участок спинного мозга, в котором происходит накопления секрета, и последующее выведение его в кровь носит название урофиза (или нейрофиза). У костистых рыб урофиз вычленяется в виде вентрального выроста спинного мозга. У хрящевых и осетровых рыб он морфологически не выражен, однако система нейросекреторных клеток имеется. Структурная организация урофиза и нейрогипофиза очень сходны между собой. Есть сведения и об участии этой системы в регуляции водно — солевого обмена: введение экстрактов урофиза карпа лабораторным крысам продемонстрировало антидиуретический эффект. (Zshikame, 1970). Показано, что урофиз влияет на уровень ионов натрия, магния и хлора в плазме крови (Fryer et. al., 1978). Вещества, выделяемые урофизом называют – уротензинами. Уротензины регулируют давление крови, сокращение гладкой мускулатуры.
8.7Половые гормоны. Половые гормоны выполняют смешанную секрецию, ястыки самок рыб вырабатывают в больших количествах гормоны эстрогены определяющих пол, половое поведение, развивают вторичные половые признаки у рыб.
Семенники самцов вырабатывают мужские половые гормоны – тестостерон, стимулирующий выработку и секрецию спермии формирующие сперматогенез, половое поведение и вторичные половые признаки.
Контрольные вопросы.
1.Обьясните понятие гормоны и инкреты.
2.Как квалифицируются гормоны по классам и функциональным признакам?
3.Поясните механизм функциональной взаимосвязи эндокринных желез.
4.Роль щитовидной железы и их гормонов.
5.Какие функциональные изменения наступают в организме рыб при дефиците или избытке инсулина?
6.Какова роль гипофиза в эндокринной системе?
7.Принципы практического использования гормонов в рыбоводстве.
Глава 9
9.1 Размножение рыб. Сохранение и увеличение рыбных запасов в естественных водоемах направлены в основном на обеспечение размножения рыб.
Размножение – это звено жизненного цикла рыбы, обеспечивающее во взаимодействии с другими звеньями воспроизводство популяции и сохранение вида. Для каждого вида рыб характерны свои специфические особенности размножения. Они обусловлены адаптацией разных видов рыб к определенным условиям размножения и развития молоди, которая дает пополнение, необходимое для сохранения вида и поддержания его численности.
У одного и того же вида рыбы плодовитость подвержена весьма большим изменениям и, в первую очередь, в связи с изменением размеров рыбы. У большинства рыб количество икринок с возрастом сначала увеличивается, а затем, с началом старения особи, уменьшается. Сильные изменения претерпевают плодовитость у одной и той же популяции в зависимости от обеспеченности пищей.
Для рыб с порционным типом икрометания, характерно одновременное созревание лишь части икры в яичнике. Промежутки между выметыванием отдельных порций икры, часто связано с продолжительностью первого этапа внешнего питания личинки. Когда личинка первой порции икрометания достигает второго этапа внешнего питания и переходит на иной вид корма, личинки следующей порции нереста только начинают внешнее питание. У большинства рыб наиболее высококачественное потомство получается от рыб среднего возраста. Молодые и очень старые дают менее жизнестойкое потомство. Первые порции спермы у самцов более высококачественные. Среди рыб одного поколения быстрее созревают особи с большей жирностью тела, однако рано созревающие рыбы обладают меньшей воспроизводительной способностью, чем поздно созревающие: плодовитость их ниже, икринки мельче, с меньшим количеством жира.
9.2 Жизненный цикл рыб. Жизненный цикл рыб распадается на различные периоды, каждый из которых характеризуется определенными морфологическими и физиологическими особенностями.
Эмбриональный период — эндогенное питание, длится от момента оплодотворения до перехода на экзогенное питание. Подразделяется на два подпериода:
— собственно эмбриональный, когда развитие происходит внутри яйцевой оболочки,
— предличиночный, когда выклюнувшиеся свободноживущие особи питаются за счет желтка.
Личиночный период — переход на активное внешнее питание с сохранением личиночных органов и признаков.
Мальковый период — по внешнему строению организм преобретает сходство со взрослым. Закладывается чешуя. Дифференциация пола, но половые органы не развиты.
Ювенильный период — усиленное развитие половых желез, но рыбы еще неполовозрелы. Начинают развиваться вторичное половые признаки.
Взрослый организм — с момента половой зрелости, делится на старческий период — замедляется или полностью прекращается рост. Рыба теряет способность размножаться.
Эмбриональный период в свою очередь разделяется на несколько стадий:
I «а» стадия — начало дробления. I стадия — дальнейшее дробление, гаструляция (обрастание желтка зародышевым слоем), до появления зародышевой полоски.
II стадия — образование зародышевой полоски. Дифференцировка туловищных миотомов. Стадия заканчивается отделением хвостовой почки от желтка.
III стадия — несформировавшегося эмбриона. Начинается от момента обособления хвоста и длится до охвата эмбрионом всего желтка.
IV стадия — сформировавшегося эмбриона. Начинается пульсация сердца и подергивание эмбриона. Длится до выхода эмбриона из икринки.
9.3 Половая система рыб. Рыбы, как правило, раздельнополы. Однако некоторые рыбы двуполы. Бывают случаи превращения одного пола в другой. Половые железы самца представлены парными семенниками (молоками), содержащими в период размножения огромное количество сперматозоидов. От семенников идут протоки, которые открываются в половое отверстие.
Половые железы самки состоят из парных (как исключение, из одного) яичников, переходящих в короткие выносящие протоки, открывающиеся в половое отверстие. В яичниках находятся икринки (яйца). Количество икринок зависит как от возраста и размеров рыб (чем старше и крупнее рыба, тем больше икры), так и от их биологических особенностей (как правило, чем меньше данный вид рыбы проявляет заботы о потомстве, тем больше икры мечут самки).
Икринки в большинстве своем очень мелкие, круглые, богатые желтком. Строение икринки (яйцеклетки) костистых рыб довольно сложное. Икринка покрыта двумя оболочками: внешняя часть бывает студенистой и липкой; внутренняя толстая оболочка иногда состоит из двух слоев – более плотного и менее плотного. Оболочки имеют отверстие – микропиле, сквозь которое внутрь икринки проникают сперматозоиды. Помимо двух оболочек имеется зародышевый диск, состоящий из протоплазмы с ядром. Запас белкового вещества сосредоточен в желтке, в определенном месте которого находится одна или много жировых капель.
Оплодотворение у подавляющего большинства рыб происходит вне тела, в воде. Процесс оплодотворения заключается в том, что сперматозоиды проникают внутрь икринки через микропиле, ядро яйцеклетки сливается с ядром сперматозоида и начинается дробление, ведущее к развитию зародыша. Зародыш развивается на питательном желтке, часть желтка остается в качестве резервного материала в виде желточного мешка у личинки.
Рыбы, как правило, раздельнополы. У костистой рыбы в верхней части брюшной полости по бокам плавательного пузыря располагаются парные половые железы. У самок половые железы представлены длинными яичниками (ovaruem), которые имеют хорошо заметную «зернистую» структуру. Задние, вытянутые отделы яичников играют роль выводных протоков и открываются непарным половым отверстием позади анального отверстия.
Половые железы самцов – длинные, гладкие, довольно плотные семенники (testis). Задние отделы семенников превратились в короткие выносящие протоки, открывающиеся общим половым отверстием позади
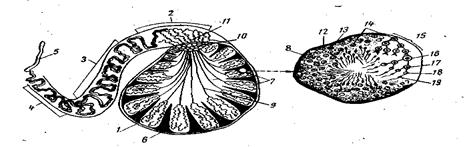
Рис.58. Схема строения семеника и поперечного разреза извитого канальца:
§
Оболчка семеника, 2-головка придатка,3-тело придатка,4-хвост придатка семеника,5-семяпровод,6-соеденительно тканные перегородки,7-извитые канальцы,8-поперечный разрез извитого канальца,9-прямые канальцы,10-сеть канальцев, семеника,11-семявыносящие канальцы,12-клетки Сертоли 13-клетки сперматогенного эпителия,14-сперматогонии, 15-последовательность размножения сперматогоний,16-сперматоцит 1-го порядка,17- сперматоцит 1-го порядка,18-сперматида,19-спермии
анального отверстия. Самцы имеют парные семенники, они лежат в виде удлиненных тел по бокам пищевода под лопастями печени. От семенников отходят тонкие семявыносящие канальцы, проходящие по брыжейке, на которой подвешен семенник.
Семявыносящие канальцы впадают в верхнюю часть почки, фактически выполняющие функцию придатков семенника. Канальцы этой части почки сливаются в семяпровод – вольфов канал. Он проходит по средней линии брюшной поверхности почки. У половозрелых самцов самая концевая часть семяпровода открывается в клоаку мочеполого сосочка. В конце каждого вольфова канала примыкает, открываясь полости мочеполового сосочка, тонкостенный половой вырост – семенной мешок. Семенные мешки являются остатками задней части мюллеровых каналов.
В семенниках образуются зрелые мужские половые клетки – сперматозоиды. Формирование мужских половых клеток-сперматозоидов, происходит в канальцах семенника. Еще не созревшие спермотозоиды через семявыносящие канальцы попадают в переднюю часть почки, здесь происходит их дозревание. Зрелые сперматозоиды скапливаются в семенных пузырьках и семенных мешочках. (Рис. 59).
Рис. 59.Схема строения сперматозоида:
При оплодотворении сокращениями стенок семенных пузырьков и семенных мешков спермотозоиды выталкиваются в клоаку самца, откуда при помощи копулятивных органов вводятся в клоаку самки.
Половых железах самок — яичниках образуются зрелые женские половые клетки- яйца (икринки). Парные яичники самок, хорошо видны по бокам пищевода. Парные яйцеводы, или мюллеровые каналы, расположены на брюшной поверхности почек. Передние концы правого или левого яйцеводов, огибая печень, соединяются в общую воронку яйцеводов, которая находится на брюшной поверхности центральной лопасти печени. В области передней части почек яйцеводы образуют небольшие расширения — скорлупывые железы, секрет которых формируют оболочку яйца. Расширенную часть каждого яйцевода называют «маткой». Матки открываются в полость клоаки в виде отверстии по бокам мочевого сосочка.
Созревшие яйца после разрыва стенки яичника выпадают и, скатываясь по поверхности печени, попадают в увеличившуюся воронку яйцеводов. Перистальтические сокращения стенки яйцеводов перемещают яйцо по направлению к матке. Для акуловых рыб характерны внутренние оплодотворения, т.е. слияние яйцеклетки со сперматозоидом происходит в верхних отделах яйцеводов.
Рис.60. Строение яйца окуня Рис.61. Спермий ильной рыбы и
§
1.Жировая капля 1.Спермий ильной рыбы
2.Кортикальные альвеолы а.Карася
3.Желточное включение б.Щуки
4.Студенистая оболочка в-г.Окуня
5.Зона радиата д-ж.Бычка
6.Плазматическая мембрана и-к.Бельдюги
7.Поверхностный слой цитоплазмы л. Форели
8.Переходная зона цитоплазмы,желток м-н.Набухшие головки
Процессы созревания половых клеток проходит в несколько стадий:
I— cтадия, у живородящих форм, яйца задерживаются в маточных отделах яйцеводов до полного формирования зародыша, а у яйцекладущих видов одетые плотной оболочкой яйца выделяются наружу. Яичники прозрачны и почти бесцветны, вдоль их проходит крупный кровеносный сосуд. В яичниках большое количество жировой ткани, хорошо различимы ооциты, которые составляют основную массу половых клеток. Вокруг ооцитов закладывается слой фолликулярных клеток.
II — стадия, это начало образования фолликулярной оболочки. Наряду с ооцитами в яичниках присутствуют также оогонии. Семенники имеют вид округлых тяжей или тонких лент от сероватого или бело- розового до багрово- красного цвета. Половые клетки у самцов представлены сперматогониями в состоянии размножения. Сперматогонии несколько раз делятся, увеличиваясь при этом в числе и уменьшаясь в размерах. Группы половых клеток носят название цист. В результате размножения сперматогонии семенники увеличиваются в размерах, теряют прозрачность, становясь мутными.
Половые железы самок хорошо развиты. Яичники занимают от трети до половины объема брюшной полости и содержат ооцит, видимые невооруженным глазом. Ооциты растут увеличиваются в объеме, накапливая в плазме питательные вещества, представленные зернами желтка и каплями жира. Этот период называется периодом трофоплазматического или большого роста. К концу периода роста ооциты достигают размеров, превышающих исходные размеры оогонии. Ооциты становятся непрозрачными, мутными и приобретают за счет жировых капель и гранул желтка окраску от светло- желтой до ярко-оранжевой. Соответственно изменяется и цвет яичников. У осетровых рыб ооциты приобретают темную окраску и на этой стадии формируются оболочки ооцита. У осетровых, кеты, окуня, сельди эта оболочка состоит из наружного и внутреннего слоев. У других рыб имеется только один слой. Эта оболочка пронизана канальцами, в которых располагаются микроворсинки ооцита.
При рассмотрении оболочки ооцита под микроскопом видна характерная радиальная исчерченность, создаваемая тончайшими порами канальцев, что и обусловило ее название — радиально – исчерченная оболочка, состоящая из межклеточного вещества. Фолликула покрыта соединительной оболочкой. У осетровых рыб на поверхности zona radiata образуется вторая оболочка, которая состоит из радиальных столбиков. Такая оболочка называется студенистой ячеистой, или сотовой оболочкой.
III-стадия зрелости яичников характеризуется наличием ооцитов с периодом трофоплазматического роста.
Семенники на III стадии зрелости значительно увеличиваются в объеме, они плотные и упругие, розовато — серого цвета. Семенные канальцы заполнены цистами. Сперматогония вступает в период роста и превращаются в сперматоциты 1-го порядка. Затем половые клетки вступают в период созревания и дважды последовательно делятся. В результате деления из каждого сперматоцита 1-го порядка возникают 2 сперматоцита 2-го порядка и 4 сперматиди, отличающиеся от исходных клеток. В процессе 2-х делений число хромосом уменьшается вдвое. Это связано с наследовательностью. Основную массу составляет ядро. Головка у сперматозоидов неодинакова по форме: шаровидная, яйцевидная, желудеобразная, палочковидная. На передней части головки находятся акросома, имеющая форму шапочки. К задней части головки примыкает средняя часть сперматозоида, переходящая в хвост, который образован осевой нитью, окруженной слоем цитоплазмы и покрыт снаружи поверхностной плазматической мембраной. Хвост осуществляет двигательные функции сперматозоида.
Сперматозоиды неподвижны в спермиальной жидкости и приобретают подвижность только в воде. Общая длина сперматозоида 50 мкл. Таким образом, в семенниках III стадии в начале присутствуют сперматогонии, сперматоциты 1-го и 2-го порядка и сперматиды, в конце появляются зрелые сперматозоиды.
IV стадия – у самок яичники крупные, достигли полного развития и занимают большую часть полости тела. Цвет яичников у разных рыб неодинаков. Ооциты закончили трофоплазматический рост и предназначены для вымета, достигли размеров икринок данного вида, вступают в период созревания. В этот период ядро ооцита смещается в микропиле. На анимальном полюсе обоих оболочек находится микропиле.
Микропиле — это канал, по которому сперматозоид проникает в яйцо и представляет собой воронкообразное углубление поверхности оболочки, которое переходит в короткий концевой каналец, открывающийся в цитоплазму на внутренней стороне радиально исчерченной оболочки. В процессе смещения ядра в ооците ядро и желток располагаются полярно. На одном полюсе находятся ядро и основная масса, на другом — желток. Затем желток и жир сливаются.
Семенники на IV стадии достигают наибольшей величины. Они имеют молочно — белый цвет. При разрезании семенника его края оплывают, закругляются, на срезе выступает густая капля спермы. На этой стадии завершается сперматогенез. В семенных канальцах семенников содержатся зрелые сперматозоиды, вышедшие из цист. Семенники содержат также и запасной фонд половых клеток, представленных сперматогониями.
V стадия – текучие половые продукты, при поднятии рыб за голову или при легком надавливании на их брюшко икра и сперма свободно вытекают из полового отверстия, завершается подготовка ооцитов к оплодотворению. У ооцитов ядро уменьшается в размерах, кариоплазма перемешивается с цитоплазмой. Затем процесс мейоза вступает в завершающую фазу, оболочка ядра растворяется и начинаются мейотические деления. Далее зрелые ооциты освобождаются от фолликулярной и соединительной оболочки. Фолликулы разрываются и ооциты выпадают в полость яичников, а у лососевых и осетровых в полость тела. Разрыв фолликул и выход икринок в полость яичников или полость тела у одних рыб протекает синхронно, у других носят растянутый характер.
У самцов на V стадии образуется семенная жидкость, которая сильно разрежает концентрацию сперматозоидов и вызывает их вытекание. Сперма имеет консистенцию молока или жидкой сметаны. Семенники мягкие на ощупь. По мере вытекания спермы размер семенников уменьшается.
VI стадия – посленерестовое состояние половых желез. Половые продукты выметаны. Яичники небольшого размера дряблые и воспаленные. Они имеют багрово – красный цвет от кровоизлияний.
Семенники уменьшились в размерах, сжались и имеют вид тонких вялых тяжей. Сперма отсутствует в семенных канальцах. Кровеносные сосуды семенников расширены, цвет семенников розоватый или буроватый. Сперматогенез продолжается во время нерестов, поэтому стадии семенников меняются.
У неоплодотворенной икры костистых рыб во всем поверхностном слое плазмы имеется множество мелких капелек, которые помещаются в вакуолях значительно большего размера. После оплодотворения икринки эти капельки выстраиваются под ее оболочку. В результате оболочка отделяется от желтка, и дальнейшее проникновение спермиев под оболочку икринки становится невозможным. Пока выделение капель не произошло, икра сохраняет способность к оплодотворению. Как только икринка оплодотворена, в ней сейчас же начинается процесс развития эмбриона.
Продолжительность инкубационного периода варьирует у одного и того же вида в зависимости от изменения различных факторов окружающей среды в частности от температуры. На развитие икры огромное влияние оказывает свет. По мере роста и развития икринок, прочность оболочки ослабевает. Выход эмбриона из оболочки обеспечивается путем ее растворения, что происходит в результате ферментативной деятельности.
Икринки снаружи покрыты оболочками, которые могут быть первичными, вторичными и третичными. Первичная — желточная или лучистая (zona radiata) оболочка, образованная самим яйцом, пронизана многочисленными порами, по которым в яйцо поступают питательные вещества во время его развития в яичнике.
Вторичная оболочка студенистая, липкая, с разнообразными выростами, предназначенными для прикрепления яиц к субстрату.
Половая зрелость у отдельных видов, особей наступает в различном возрасте. Большинство карповых, окуневых и лососевых рыб достигают половой зрелости в возрасте 2-6 лет, а осетровые становятся половозрелыми в возрасте 6-12 лет. При этом необходимо отметить, что половозрелость у самцов, как правило, наступает на 1-2 года раньше, чем у самок. Под влиянием факторов внешней среды (прежде всего температуры и корма) развитие половых клеток может ускоряться или замедляться, а в некоторых случаях даже приостанавливаться. Так, например, лещ, обитающий в наших южных водоемах, становится в массе половозрелым на 3-4 м году, в водоемах центральных районов страны на 4-5 м году, в наших северных водоемах со сравнительной низкой годовой температурой половозрелость у леща наступает на 5-7м году жизни. Аналогичную картину по скорости созревания половых клеток можно наблюдать и у других видов рыб.
Разнообразен и интересен процесс размножения рыбы. Такие, как сом, щука, могут в течение своей жизни нерестовать много раз, а дальневосточные лососи: кета, семга, горбуша — нерестуют лишь один раз в жизни и после первого же нереста погибают от физического истощения.
Недостаточное питание рыб задерживает созревание их половых клеток,а полноценное питание, полностью обеспечивающее организм в питательных веществах, ускоряет их созревание. Известно также, что уменьшение численности популяции рыбы под влиянием промысла приводит к повышению обеспеченности пищей, а это отражается на ускорении полового созревания особей.
Сдвиги наступления половой зрелости у рыб могут происходить под действием факторов внешней среды. Например, волжская белуга достигает половой зрелости в возрасте от 10 до 18 лет.
Стадии зрелости половых желез можно определить при помощи разработанных для каждой группы рыб, так называемых шкал зрелости. Так, для карповых и окуневых существуют шкалы С.Н.Кулаева и В.А.Мейена, для осетровых – шкалы А.Я.Недоливина, А.В. Лукина и И.Н.Молчановой. Каждая из этих шкал показывает специфические особенности процесса созревания половых продуктов только у данной группы рыб. О.Ф.Сакун и Н.А. Буцкая разработали две универсальные шкалы для всех наших промысловых групп рыб. Первая шкала отражает общий ход созревания половых продуктов у самок, а вторая – у самцов. Эти шкалы позволяют определить стадии зрелости половых желез по их внешнему виду и гистологическому строению.
Для оценки степени зрелости половых продуктов у рыб применяется шестибалльная шкала зрелости.
I стадия – ювенильная, неполовозрелые молодые особи. Половые железы имеют вид тонких прозрачных желтоватого или розоватого цвета тяжей, прилегающих к стенкам полости тела. Половые клетки у самок представлены или только оогониями, или оогониями и молодыми ооцитами периода протоплазматического роста. Оогонии – это первичные половые клетки, которые образуются у самок рыб из зачаткового эпителия в течение всей жизни. Оогонии развиваются и длятся, что приводит к увеличению их численности. И этот период называется периодом деления оогоний, затем наблюдается их рост, и растущие клетки называются ооцитом.
Половые клетки самцов представлены сперматогониями. Сперматогонии – это первичные крупные половые клетки в семенниках, округлой формы с большим ядром, окруженным широким слоем цитоплазмы, которые образуются у самцов рыб из перитонеального эпителия.
II- стадия — покоя. Половые продукты или еще не начали развиваться, или были выметаны. Икринки очень мелкие и видны только под лупой. Яичники прозрачны и бесцветны, вдоль них проходит крупный кровеносный сосуд. Гонады занимают 1/2 полости тела.
III стадия — созревания. В икринках начанается накопление желтка, и переход к этой стадии свидетельствует о наступлении полового созревания. Икринки заметны невооруженным глазом. Гонады занимают 2/3 полости тела, половые железы хорошо развиты.
IV стадия — зрелости. Рост икринок закончился. Яичники и семенники крупные, достигли максимальных объемов, массы и полного развития и занимают большую часть полости тела. При надавливании на брюшко самца появляется капля спермы. Легкое надавливание на брюшко самки не приводит к вытеканию икры.
V стадия — текучести. Гонады занимают почти всю полость тела. При легком надавливании на брюшко икра и молоки вытекают наружу.
VI стадия — выбоя. Икра выметана, молоки вытекли, посленерестовое состояние половых желез.
Развитие женских половых клеток (оогенез) и мужских половых клеток (сперматогенез) – длительный и сложный процесс. Каждая половая клетка, прежде чем она окончательно созреет, должна обязательно пройти в своем развитии ряд стадий. При этом следует различать два процесса; 1 – период до достижения половой зрелости, начиная от возникновения первичных половых клеток и кончая образованием зрелых половых продуктов; 2 – периодическое созревание определенной части половых продуктов в течение межнерестового периода (после достижения половой зрелости). Первый процесс более длительный, чем второй, и у разных видов рыб занимает разное время. Периодическое же созревание половых продуктов у половозрелых особей требует меньшего времени, но продолжительность его у разных видов рыб различна. Так, сазан, лещ, судак и многие другие виды рыб размножаются, как правило, ежегодно, а осетровые рыбы лишь через 3-5 лет, а отдельные особи – через 9 лет. Исключение составляют тихоокеанские лососи, которые после размножения погибают.
§
Миграциями называют закономерные направленные перемещения животных в пространстве. В отличие от перемещений тип миграции характеризуют следующие особенности:
1. Строгая сезонность
2. Множественная перестройка физиологических систем организма в соответствии со специфическими задачами миграции.
3. Массовость. Мигрируют не отдельные особи, а целые популяции. Годичный цикл рыб в наиболее полном виде включает три типа миграции: нерестовые (миграции в место размножения), нагульные (кормовые) и зимовальные. Эти типы миграции не у всех видов четко выражены. В ряде случаев места нагула и зимовки или размножения и нагула совмещаются, соответственно выпадает одна из форм миграций.
По направленности миграции рыб принято делить на анадромные (из моря к берегам и далее вверх по рекам) и катадромные (в обратном направлении).
Помимо внешних условий, состояние миграции зависит от физиологического состояния организма. Так, зимовальные миграции стимулируются определенной степенью упитанности и жироотложения.
Нерестовая миграция обычно подготавливается определенной степенью зрелости половых продуктов и специфическим состоянием «гормонального фона», организма. У проходных рыб стимул к нерестовой миграции возникает после длительного (от 1 до 15-16 лет) периода морской жизни. У рыб наиболее общий показатель, стимулирующий миграцию — высокая жирность. По мере приближения к местам нереста жировые запасы уменьшаются, что отражает высокий уровень затрат энергии на миграцию и созревание половых продуктов. Так, жирность каспийской миноги Caspiomyzon осенью при входе в Волгу составляет 30-35%, а вблизи нерестилищ падает до 1-2%;
Важной характеристикой состояния мигрирующих рыб является функциональная активность гипоталамической нейросекреторной системы и гипофиза. Во время пресноводной миграции регистрируется высокий уровень продукции гонадотропинов. Наблюдается усиление тиреотропной функции гипофиза и прогрессивное развитие щитовидной железы при движении вверх по реке. Последнее связано с усиленными затратами энергии. Показано, например, что собственная скорость движения кеты Oncorhynchus keta по Амуру достигает 30-50 кмсут. (с учетом скорости течения более 100 кмсут), что соответствует расход энергии порядка 100-118 тыс. кдж на 1 кг массы в сутки (Шмидт, 1947). По прибытии на нерестилища активность щитовидной железы снижается.
Факторы, стимулирующие начало миграции, могут быть различными. Это повышение температуры воды в реках и начало поводка. Фотопериод внешние стимулы действуют только на «подготовленный» организм. Состояние физиологической готовности к миграции предшествует началу внешней стимуляции.
Основными связующими звеньями между сезонным ходом внешних факторов и внутренней физиологической перестройкой является нейросекреторная и эндокринная системы. (Рис.62)

При этом реализуются специфические формы поведения, стимулируется ориентированная миграционная активность, перестраивается система осморегуляции, организм становится чувствительным к влиянию внешних факторов среды (температура, соленость, уровень воды, течение и пр.)
Катадромные миграции молоди могут быть разных типов. Лососевые мигрируют к морю активно, что определяется закономерными изменениями обмена осморегуляции и других процессов. Перечисленные изменения регулируются на уровне гипоталамо-гипофизарной системы и стимулируемых этой системой эндокринных комплексов. У осетровых миграция к морю имеет пассивный характер.
9.5 Нерест и оплодотворение. Нерест — самый ответственный этап. Нерест является классическим годовым биоритмом рыб. У каждого вида рыб он наступает при определенном наборе условий, хотя температурные режимы для одного и того же вида рыб могут различаться в зависимости от местоположения водоема. Функции размножения живых организмов носят циклический характер. Каждый цикл состоит из отдельных фаз, связанных с деятельностью гормонов и раздражителей внешней среды.
Нерест — самый ответственный этап. Нерест является классическим годовым биоритмом рыб. У каждого вида рыб он наступает при определенном наборе условий, хотя температурные режимы для одного и того же вида рыб могут различаться в зависимости от местоположения водоема. Функции размножения живых организмов носят циклический характер. Каждый цикл состоит из отдельных фаз, связанных с деятельностью гормонов и раздражителей внешней среды.
Рыбы, как и теплокровные животные, привлекают особей другого пола посредством выделения ими специальных химических веществ — феромонов, а также с помощью сигнализации, определенных поз, телодвижений, брачной окраски и т.д. Основными составляющими предпосылок, дающих «команду» к нересту, являются температурный фактор, кислородный режим, наличие субстрата для откладывания половых продуктов, наличие партнера. При отсутствии любого из них рыбы на нерест не пойдут, икра может рассосаться.
Икрометание бывает единовременным, когда рыбы выметывают икру в течение нескольких минут, часов, дней (щука, окунь, плотва и другие), или порционным, когда рыбы мечут икру отдельными порциями с интервалами от нескольких дней до 2-3 недель — линь, густера, карась и другие. Одни и те же виды рыб в различных климатических условиях мечут икру по-разному. Лещ, например, в южных районах нерестится порционно, а в северных — единовременно. Рыбам, видимо, нужно до холодов восстановить силы, а главное, требуется определенное время для того, что молодь успела до зимы окрепнуть.
 Многократному икрометанию рыбы способствуют периоды, связанные с определенными фазами Луны, рыбы ждут эту фазу в течение лунного месяца. Если условия для нереста при нужной фазе были неподходящими, нереста может и не произойти. Может быть, по этой причине нерест серебристых карасей длится с конца мая месяца до августа. Рыбы нерестятся не всем своим сообществом сразу, а поочередно, по возрастным группам и в определении сроков нереста могут оказать фенологические признаки природы.
Многократному икрометанию рыбы способствуют периоды, связанные с определенными фазами Луны, рыбы ждут эту фазу в течение лунного месяца. Если условия для нереста при нужной фазе были неподходящими, нереста может и не произойти. Может быть, по этой причине нерест серебристых карасей длится с конца мая месяца до августа. Рыбы нерестятся не всем своим сообществом сразу, а поочередно, по возрастным группам и в определении сроков нереста могут оказать фенологические признаки природы.
Достигнув половой зрелости, рыбы совершают нерестовые миграции, то есть они перемещаются от мест нагула или зимовки к местам икрометания, где завершается у них созревание половых клеток и происходит нерест. Морские рыбы постоянно живут и размножаются в море. Они размножаются в основном в тех же местах, где и постоянно обитают, или же вблизи от них. Проходные рыбы живут в море, а размножаются в реках, по которым большинство из них проходит сотни километров, преодолевая сильные течения, водопады, пороги и препятствия, встречающиеся на их пути к местам икрометания. Проходные рыбы -это те рыбы, которые живут в реках и озерах, а для размножения выходят в море.
В нерестовый период у самок рыб, у которых половые железы находятся в IV стадии зрелости, под воздействием гонадотропного гормона гипофиза завершается оогенез и происходит овуляция. Овуляция у рыб осуществляется только при определенных условиях внешней среды, как температура, гидрохимический режим, наличие или отсутствие течения воды, наличие самцов, для многих рыб нерестовый субстрат. После овуляции происходит брачный акт, который называется нерестом, а то место, где он совершается, — нерестилищем.
Место нереста является приспособлением рыб к наиболее благоприятным условиям для эмбрионального и постэмбрионального развития. В связи с этим С.Г. Крыжановский разделил всех на следующие экологические группы;
— Литофилы размножаются на каменистых, гравийных и твердых глинистых грунтах рек с быстрым течением и олиготрофных озер, в которых имеются благоприятные условия для дыхания зародышей. К этой группе относятся лососи, сиги, осетровые и некоторые карповые рыбы.
— Фитофилы размножаются в реках, озерах, водохранилищах и прудах, откладывая икру на растительный субстрат в стоячей или слабопроточной воде. К этой группе относятся лещ, сазан, судак, щука, окунь, плотва, карась и др.
— Псаммофилы размножаются в реках и озерах с песчаными группами и при наличии благоприятных условии для дыхания зародышей. К этой группе принадлежат пескари и некоторые гольцы.
— Пелагофилы размножаются в реках и озерах. К этой группе относятся проходные сельди, чехонь, белый амур, белый и пестрый толстолобики и др.
— Остракофилы размножаются в реках, откладывая икру в жаберную полость двухстворчатых моллюсков. К этой группе относятся горчаки.
Продолжительность нереста. Нерест рыб разных пород происходит не в одно время. Обычно нерест той или иной породы рыбы начинается когда вода достигнет наилучшей для нее температуры, т. е. когда икра рыбы созреет и приобретет способность течь. Например лосось севера — семга нерестится в октябре — ноябре при температуре воды 2—6°, щука при 7— 10°, а сом при 18—22°. Поэтому верным показателем начала нереста следует считать температуру воды. Также нельзя обобщать сроки для любой местности и каждой породы рыбы, например окунь в реке Дон нерестится в феврале — марте, в Московской области—в апреле—мае, в водоемах Архангельской области или в реке Енисей — в июне. Продолжительность нереста у каждой породы рыб разная, например; у окуня нерест продолжается 3—5 дней, он откладывает икру сразу, а уклейка — в несколько приемов, растягивая свой нерест на длительное время. Нерест вследствие неблагоприятных условий, например, понижения температуры воды может, начавшись, прерваться.
Каждому виду рыб свойственна определенная температура воды, при которой может произойти нерест, и требуются для этого соответствующие нерестилища. У большинства рыб самцы первыми подходят к нерестилищам. Затем на местах нереста появляются самки. Соотношение самок и самцов на нерестилищах обычно один к одному. Однако это соотношение не является постоянным, может изменяться у одной и той же популяции по годам.
Во время нереста самки выметывают икру, а самцы осеменяют ее. Соприкосновение спермы с икрой называется осеменением.
Самки после вымета икры уходят с нерестилищ. У большинства рыб самцы после осеменения икры остаются на нерестилищах. Они принимают участие в нересте с другими самками, которые подходят на нерестилища. В течение нерестового периода один самец может осеменять икру нескольких самок.
Продолжительность периода икрометания отдельной особи зависит от особенностей созревания половых клеток у данного вида рыб. Если рыба обладает единовременным типом созревания ооцитов, то и нерест разовый, кратковременный. Так, вобла выметывает икру за одно утро. Продолжительность нереста одного вида рыб зависит от типа икрометания, от структуры нерестового стада, от наличия в популяции экологических групп, от температуры воды. Экологические особенности размножения рыб необходимо учитывать при рыборазведении и строительстве искусственных нерестилищ.
Время нереста у различных видов рыб весьма разнообразно и приурочено к различным сезонам года, поэтому существуют внутривидовые биологические группы рыб. Наших промысловых рыб, у которых нерест происходит в различные сезоны, разделяют на две группы:
1.Весенне — летненерестующие: щука, сом, судак, вобла, сазан, лещ, рыбец, кутум, осетр, севрюга, белуга, шип и многие другие виды:
2.Осенне – зимненерестующие: лососи, сиги, белорыбица, налим и многие другие виды.
Проходные рыбы идут на нерест в реки в различное время года и с различной степенью зрелости половых желез. Одни из этих рыб нерестятся вскоре после захода в реку, другие только на следующий год. Наблюдая такое явление, Л.С.Берг пришел к заключению, что у некоторых лососевых и осетровых рыб существуют яровые и озимые расы. Яровые рыбы мигрируют из моря в реки, где они мечут икру в этом же году. Озимые идут из моря в реки, где они нерестятся только на следующий год.
Н.А.Гербильский, Б.Н.Казанский и И.А.Баранникова, изучив особенности размножения куринского и волжского осетра, установили наличие биологических групп внутри их популяции, развив тем самым учение Л.С.Берга о сезонных расах. Так, осетр, мигрирующий из Каспия в Волгу, имеет четыре биологических группы: ранний яровой осетр, поздний яровой осетр, озимый осетр летнего хода и озимый осетр осеннего хода.
Кроме куринского и волжского осетра, внутрипопуляционная дифференциация существует у осетра и в ряде других рек. Биологические группы имеются также внутри популяции севрюги и белуги в реках Каспийского и Азовского бассейнов.
Места нереста озимых рыб расположены в верхнем течении рек, а яровых рыб – в нижнем. Такая внутривидовая дифференциация проходных рыб в отношении мест нереста также носит приспособительный характер и позволяет использовать нерестилища, расположенные в различных точках реки на очень большом ее протяжении благоприятные условия для выживания потомства. После нереста производители проходных рыб и их молодь мигрирует из рек в море. Во время нереста рыб происходит осеменение икры. Сперматозоид проникает внутрь икринки через отверстие в ее оболочке – микропиля. При слиянии женского и мужского ядер (пронуклеусов) происходит оплодотворение, которое дает начало развитию эмбриона (зародыша). С этого момента начинается развитие организма рыбы.
В процессе онтогенеза рыб выделяют следующие периоды, эмбриональный, личиночный, мальковый и половой зрелости. Каждый период характеризуется своей качественной спецификой. Для эмбрионального периода характерно эндогенное питание желтком желточного мешка. Для личиночного периода характерно питание внешней пищей. Они имеют временные личиночные органы. В мальковый период эти органы исчезают.
Группа последовательных этапов развития, объединенных одинаковой качественной спецификой, составляет период развития. Теоретические основы этапности развития рыб были разработаны В.В. Васнецовым, который сформулировал основные положения этой теории. Они сводятся у тому, что в течение различных периодов онтогенеза (эмбрионального, личиночного, малькового и половой зрелости) развитие рыб идет не только постепенно и непрерывно, но и прерывисто, скачкообразно.
Эти морфологические изменения неразрывно связаны с изменениями биологических особенностей рыб. Между изменениями происходят постепенный рост, медленные качественные особенности, организация и его отношения с внешней средой сохраняются неизменными. Такие интервалы относительной стабильности развития рыб между двумя скачкообразными изменениями называются этапами. Каждый этап развития рыб характеризуется определенными особенностями строения организма, биологическими свойствами и требованиями к условиям внешней среды. При отсутствии условий рыбы задерживается на том или ином этапе, замедляют или прекращают рост и погибают. Развитие рыб есть последовательная смена приспособлений их организма к внешней среде в течение определенных этапов развития.
9.6 Эмбриональный и личиночный периоды развития.Оплодотворенная икринка – это уже развивающийся зародыш, который проходит ряд этапов развития. Так, на первом этапе оплодотворения, после проникновения сперматозоида в икринку, наблюдаются сложные преобразования через 2-15 мин. Этот этап заканчивается соединением женского и мужского пронуклеусов. Этап дробления — начинается с момента появления на икринках первой борозды, которая образует два бластомера. Каждая икринка последовательно делится на все большее количество бластомеров, а в конце достигает стадий бластулы, начинают формироваться тело зародыша и его желточный мешок.
На следующем этапе развития в зародыше происходит нейруляция. Образуется нервная трубка, которая представляет собой зачаток центральной нервной системы.
В дальнейшем у зародыша изменяется форма тела, обособляется хвостовой отдел, формируются отделы головного мозга, закладываются органы чувств и железы вылупления.
У зародышей начинают формироваться пищеварительная и выделительная системы, образуются сердце и кровеносные сосуды, закладываются жаберные карманы, появляется зачаток печени, образуются мышечные волокна, клетки крови и начинается вакуолизация клеток, хорда зародыша расчленяется на отдельные сегменты, и на нем появляется плавниковая складка.
Зародыш имеет функционирующее сердце и кровообращение, а также нервную систему и способен отвечать на внешние раздражения. В это время зародыши освобождаются от оболочки, а вылупившиеся предличинки завершают свое развитие и становятся личинками. Огромное влияние на развитие зародыша (эмбриогенез) оказывают температура воды, газовый режим, рН, соленость, осмотическое давление и освещенность.
Эмбриогенез у каждого вида рыб протекает в определенных температурных границах, при этом различают температурный оптимум, температурный порог и температурный максимум. Зародыш у рыб с весенне-летним нерестом развивается в течение нескольких суток, у рыб с осенне – зимним нерестом в течение нескольких месяцев. Температура, при которой эмбриогенез протекает наиболее нормально, называется температурным оптимумом. Повышение температуры ускоряет эмбриогенез, а понижение замедляет этот процесс. Температура, ниже которой эмбриогенез не происходит, называется температурным порогом, однако пороговая температура не приводит к гибели зародышей. Температура, выше которой эмбриогенез прекращается и зародыш погибает, называется температурным максимумом.(Рис.63-64).
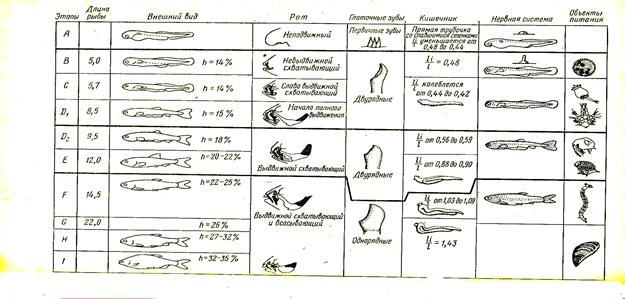
Рис.64. Изменение формы тела, некоторых органов и характера пищи у каспийской воблы.
9.7 Получение зрелых производителей.Зрелые производители — это рыбы, у которых половые продукты (икра или сперма) пригодны для оплодотворения. Зрелые самки обычно имеют мягкое брюшко, при самом незначительном надавливании на которое из генитального отверстия выделяются икринки. У зрелых самцов при легком нажиме на брюшко вытекает сперма. Получение зрелых производителей относится к важнейшему звену искусственного рыборазведения.
Заготовку производителей осуществляют рыбоводные заводы, временные рыбоводные пункты и рыбоводные нерестово-выростные хозяйства. Сроки заготовки производителей обычно приурочивают к их массовому нерестовому ходу из моря реки.
Производителей ловят в реках плавными сетями, неводами, заколами-ловушками. Рыбоводы тщательно осматривают пойманных рыб и выбирают зрелых производителей. Мелких и травмированных производителей они отбраковывают. Нередко лишь незначительное количество производителей имеет половые продукты, которые можно оплодотворить. Много производителей вылавливают незрелыми. Незрелых рыб в прорезях и на автомашинах доставляют на рыбоводный завод или на рыбоводный пункт и выдерживают до полного созревания половых продуктов. Выдерживают производителей в садках или бассейнах, в которых созданы условия, близкие естественным. Такой метод выдерживания производителей называют экологическим.
Кроме экологического метода получения зрелых производителей, существует метод, который основан на физиологическом воздействии, вводимых в организм рыбы веществ, ускоряющих процесс созревания половых продуктов у самок и самцов, имеющих IV (не ниже) стадию зрелости. Такими веществами являются гормоны гипофиза рыб. При внутримышечных инъекциях производителям суспензии гипофиза рыб гормоны поступают в кровь, разносятся по всему организму, активизируют органы, регулирующие половой процесс, и производители быстро созревают.
Применяемый в рыбоводстве для получения зрелых производителей метод гипофизарных инъекций разработал Н. Л. Гербильский. Метод гипофизарных инъекций, дающий возможность получать за 6-48 ч. зрелых производителей, в рыбоводстве принято называть физиологическим методом стимулирования созревания половых продуктов у рыб.
Для инъецирования производителей можно применять гипофизы, взятые от любых видов рыб в пределах одного семейства. Однако лучше применять гипофизы данного вида рыб. Гормоны гипофиза леща и сазана стимулируют процесс созревания половых продуктов у многих видов рыб, относящихся к различным семействам. При промышленном разведении рыб гипофизы осетровых обычно используют для инъецирования осетровых, гипофизы судака – для инъецирования судака, гипофизы сазана и леща – для инъецирования карповых рыб.
Активность гипофиза достигает максимума непосредственно перед нерестом. После нереста содержание гормонов в гипофизе резко сокращается и снова начинает увеличиваться, спустя 3-5месяцев. Поэтому заготовку гипофизов весеннее — нерестующих рыб следует проводить перед нерестом, осенью или зимой. Гипофизы от неполовозрелых рыб не заготовляют, так как они не содержат необходимого количества гонадотропного гормона.
Гипофизы заготовляют на рыбокомбинатах или непосредственно на рыбных промыслах. У только что пойманных рыб вырезают кусочки мозга и извлекают из них гипофизы, которые помещают в стеклянные банки с притертыми пробками и для обезжиривания и обезвоживания заливают безводным ацетоном. Объем ацетона должен быть в 10-15раз больше объема гипофизов. Через 12 ч. ацетон из банок сливают и наливают новую порцию его в том же объеме. Гипофизы выдерживают во второй порции ацетона еще на 6-8ч. Затем ацетон сливают из банок, а гипофизы раскладывают на бумаге и высушивают при комнатной температуре. Высушенные гипофизы хранят в тех же банках с притертыми пробками или в герметически закупоренных пробирках или в бюксах.
Перед инъецированием рыб рассчитанную дозу гипофизов растирают в фарфоровой ступке в порошок, который тщательно перемешивают в физиологическом растворе (0,5г. поваренной соли, растворенной в 1л. дистиллированной воды). Полученную суспензию гипофиза при помощи шприца вводят в спинные мышцы производителей. Доза гипофизарных инъекций зависит от вида рыбы, веса производителей, степени зрелости их половых продуктов, а также от факторов внешней среды (температуры и т. д.) действующих на организм рыбы. В настоящее время для стимулирования созревания половых продуктов у судака применяют также внутримышечные инъекции медицинского препарата хлориоганина.
9.8 Взятие зрелой икры и спермы у производителей рыб. Икру и сперму берут у зрелых производителей рыб при ровном рассеянном свете, отсутствии прямых солнечных лучей или яркого электрического освещения. Температура воздуха должна быть близкой к температуре воды.
Зрелые половые продукты берут у производителей тремя способами: путем отцеживания, вскрытия брюшной полости рыбы и комбинированным способом.
Способ отцеживания заключается в том,что перед взятием икры голову и хвостовой стебель самки обертывают влажным полотенцем или марлей. Если самка небольшая, икру отцеживает 1человек. Он прижимает голову рыбы локтем левой руки к телу, а кистью этой руки держит хвостовой стебель в таком положении, чтобы генитальное отверстие находилось над краем сухой и чистой посуды. При этом самка изгибается брюшной стороной наружу. От давления стенок брюшной полости часть икры выделяется из генитального отверстия, попадает на край подставленной посуды и стекает на дно. Затем, осторожно сдавив большим и указательным пальцами правой руки стенки брюшка самки (у грудных плавников) с обеих сторон, он сгоняет оставшуюся икру к генитальному отверстию. Отцеживание икры продолжают до тех пор, пока не прекратится выделение свободных икринок.
Если самка крупная, то икру отцеживают 2 человека: один держит голову, другой держит над краем посуды хвостовой стебель и одновременно свободной рукой отцеживает икру.
Таким же способом берут у производителей сперму. Зрелого самца держат над посудой и рукой слегка массируют его брюшко до тех пор, пока из генитального отверстия и начнет вытекать сперма. Сперму у мелких самцов можно брать, изогнув дугой их туловище (брюшком наружу), у крупных самцов с помощью резинового щупа, вставленного в генитальное отверстие. В этом случае, чтобы рыба не билась, ее необходимо поместить в станок. Когда появятся первые сгустки молок, отцеживание прекращают.
Сперма часто созревает отдельными порциями, поэтому ее можно брать у самцов несколько раз. Для этого самцов после каждого отцеживания сажают в специальные садки для выдерживания, при котором созревает очередная порция спермы.
Способ вскрытия. Самку убивают ударом колотушки по голове и обескровливают, делая ножом глубокий надрез на затылке или надрезав жабры, или перерезав хвостовую артерию. Для того чтобы вытекло больше крови, которая может попасть в полость тела и в икру, рыбу подвешивают на блок. Когда сток крови полностью прекратится, рыбу обмывают водой, вытирают полотенцем и ножом разрезают брюшко от генитального отверстия до передней его части на 7-15см. Затем из самки извлекают в заранее подставленную посуду всю икру.
Комковатую незрелую икру осторожно отделяют птичьим пером от зрелой икры. Попавшую в посуду воду быстро сливают, чтобы не дать возможности икринкам набухнуть.
Комбинированный способ. При этом способе основную часть половых продуктов берут у рыбы способом отцеживания, а оставшуюся часть их путем вскрытия брюшной полости.
9.9 Методы учета количества икры.Объемный метод. При учете икры по объемному методу обычно используют мерные кружки емкостью 0,5-1,0л. и мерные стаканчики на 1-5см. Первоначально кружками измеряют объем всего количества икры. Затем заполняют икрой стаканчик и просчитывают содержание икринок в нем. Заполнение стаканчика икринками и их подсчет повторяют 3 раза для установления в определенном объеме стаканчика, устанавливают количество икринок, находящихся во всем измеренном объеме взятой от самок икры.
Весовой метод. При этом методе первоначально взвешивают все количество взятой от самок икры: затем берут 2-3 небольшие порции икры, взвешивают их, поштучно просчитывают количество икринок в одном грамме. Зная количество икринок в 1грамме икры, устанавливают количество всей икры.
Плодовитость. Воспроизводительная способность рыб зависит от их абсолютной плодовитости. Абсолютная плодовитость – это количество находящихся в яичниках (гонадах) самки икринок, которые могут быть выметаны в нерестовой период года. Абсолютную плодовитость обычно определяют весовым методом учета икры. Для этого у взвешенной и измеренной рыбы вынимают гонады, взвешивают икру, берут порцию икры от 0,2 до 20г. (в зависимости от размеров икринок) и подсчитывают количество икринок в ней, поэтому берут не менее трех порций икры из различных участков и производят подсчет икринок. Затем определяют среднее количество икринок в 1 г. Это дает возможность установить абсолютную плодовитость, значительно труднее определять абсолютную плодовитость этим методом при порционном икрометании.
Относительная плодовитость – это количество икринок, приходящееся на единицу веса (1г. 1кг.) или длины (1см.) самки.
Рабочая плодовитость – это количество зрелых икринок, выметанное самкой в данном году в условиях рыбоводного хозяйства или взятое из ее ястыков для искусственного осеменения.
Размеры икринок у рыб разных видов сильно отличаются. Наиболее плодовитые рыбы имеют наиболее мелкую икру, и наоборот. Так, диаметр икринки сазана 1,5-2 мм, щуку 2,5-3, севрюги 2,4-3,2, осетра 2,8-3,8, белуги 3,3-4, лосося 5-7мм. У самок одного и того же вида рыб величина абсолютной плодовитости повышается с увеличением их размеров, массы возраста (обычно до начала старения) и улучшением условий их обитания, в частности питания.
Качество спермы. Качество спермы оценивают по ее концентрации и активности. Концентрация спермы – это количество сперматозоидов в единице объема семенной жидкости. Количество сперматозоидов устанавливают под микроскопом в счетной камере.
Активность спермы – это продолжительность поступательных движений сперматозоидов в воде, которую определяют с помощью микроскопа. Для этого на предметное стекло наносят каплю спермы, и устанавливают время от начала до конца поступательных движений сперматозоидов.
Активность спермы некоторых рыб
Осетровые 15-20 мин. Сазан 1,5-3,0 мин.
Семга 45 сек. Лещ 2,5-3,0 мин.
Форель 23-60 сек. Рыбец 30-60 сек.
Сиги 1 мин. Плотва 2,4 мин.
Ряпушка 3,5мин. Линь 11,5мин.
Сельдь волжская 1мин. Окунь 1,5-2,0мин.
Густая сперма отличается высокой концентрацией. Чем выше концентрация спермы данного вида рыб, тем выше активность, а также осеменяющая и оплодотворяющая способность ее.
Опытные рыбоводы обычно оценивают качество спермы визуально — по ее внешнему виду. Хорошая сперма имеет консистенцию жидкой сметаны и слегка желтоватый оттенок (у осетровых) и чисто белый цвет. Средняя по качеству сперма имеет консистенцию сливок и белый цвет. Плохая сперма – жидкая, имеющая вид разбавленного молока голубоватого оттенка. Для осеменения икры каждой самки используют смешанную доброкачественную сперму от 3-4 самцов.
Осеменение икры. Осеменение – это соприкосновение спермы с икрой. Попавшей на икринку сперматозоид по микрокапиллярному канальцу (микропиле) проникает через ее оболочку внутрь икринки, где происходит слияние женской и мужской клеток, или оплодотворение, дающее начало новой клетке – зиготе, которая делится и превращается в многоклеточный зародыш (эмбрион).
Способы искусственного осеменения. Основная задача искусственного осеменения – создание условий, обеспечивающих проникновение сперматозоида в каждую икринку. В рыбоводстве применяют 3 способа искусственного осеменения: сухой, полусухой и мокрый.
Сухой способ. В эмалированный таз с икрой добавляют сперму, после этого перемешивают. Затем в таз наливают воду и снова перемешивают половые продукты.
Полусухой способ. В эмалированный таз с икрой приливают сперму, разведенную водой непосредственно перед осеменением, и сразу же приступают к перемешиванию половых продуктов.
Мокрый способ. В эмалированный таз с икрой наливают воду, затем вносят сперму и тут же производят перемешивание. К этому способу относится и такое осеменение, когда икру и сперму одновременно вносят в эмалированный таз с водой и перемешивают.
В зависимости от вида рыб для осеменения 1 кг. икры любым из указанных 3 способов требуется от 0,5 до 10,0 см. спермы и от 0,15 до 2,0 л. воды. После перемешивания в воде половых продуктов икра оплодотворяется. Время, затрачиваемое на перемешивание в воде половых продуктов до момента оплодотворения, составляет 1-5 минут и не зависит ни от способа осеменения икры, ни от количества осеменяемой икры, а имеет прямую связь с продолжительностью поступательных движений сперматозоидов разных видов рыб.
Подготовка икры к инкубации. Перед тем, как поместить оплодотворенную икру в инкубационные аппараты, ее тщательно отмывают от остатков спермы, слизи, полостной жидкости и при необходимости ликвидируют ее клейкость.
Отмывка неклейкой и слабоклейкой икры не вызывает особых затруднений. Для этого в таз с оплодотворенной икрой наливают чистую воду (объем воды должен в 3 раза превышать объем икры) и начинают осторожно круговыми движениями руки или пучком птичьих перьев перемешивать. Затем несколько раз сливают помутневшую воду и наливают свежую. Когда сливная вода станет чистой, отмывку икры прекращают. На рыбоводных заводах икру обычно отмывают в проточной воде, поступающей в таз из резинового шланга и вытекающей через край.
Значительно труднее готовить к инкубации клейкую икру. Для отмывки такой икры берут воду, в которую предварительно вносят мелкий речной ил или измельченный в пудру мел. На отмывку 1кг. икры требуется 4л воды и 0,5л густой взвеси ила. Перемешивание икры с илом при периодическом сливаний мутной воды и добавлении чистой производится в течение 40-50 мин. После того как ил будет полностью удален из таза, вода становится прозрачной, а икра теряет способность приклеиваться.
Икра во время отмывки частично набухает. Для дальнейшего набухания икру оставляют в тех же тазах с проточной водой или помещают в сетчатые ящики, установленные в реке, или закладывают непосредственно в инкубационный аппарат, в котором осуществляется постоянная смена воды.
Если инкубацию икры проводят на субстрате в приклеенном состоянии, то осеменение икры, отмывку ее от остатков спермы, слизи и полостной жидкости лучше проводить в стерильной воде и без добавления ила или мела. Стерилизация воды осуществляется при помощи бактерицидных установок. Продолжительность отмывки этой икры в воде обычно не более 2мин. За это время клейкое вещество еще не появляется на оболочках икринок. Поэтому их свободно рассеивают на субстрате без образования комков. В дальнейшем икринки выделяют клейкое вещество и приклеиваются к субстрату
Контрольные вопросы.
1.Какими морфологическими и физиологическими особенностями
характеризуется жизненный цикл рыб?
2.Морфологические и функциональные особенности органов размножения
костистых и хрящевых рыб.
3.Функции половых органов самцов и самок.
4.Охарактеризуйте стадии созревания половых клеток самок и самцов.
5.В каком возрасте наступает половая зрелость у отдельных особей рыб и
каким образом оценивается степень зрелости половых продуктов?
6.Миграция,факторы, стимулирующие начало миграции.
7.Нерест,факторы, стимулирующие начало нереста,продолжительность
нереста
8.Механизм оплодотворения,плодовитость.
9. Каким способом искусственно осеменяют икру?
10.Обьясните суть метода Н.Л.Гербильского в рыбоводстве.
11.Технология и методы взятия, учета количества зрелой икры и спермы у
рыб
12.Способы оценки качества спермы
ГЛАВА 10
§
10.1Механизм движения рыб и роль плавников Механическое движение занимает важное место в жизненных процессах. Рыба перемещается в воде — плавает, прокачивает воду сквозь жаберный аппарат, сердце проталкивает кровь по сосудам, кишечник проталкивает пищу. Имеются и другие, менее заметные, но тоже очень важные формы механического движения — изменяется просвет кровеносных сосудов зрачка глаз, сокращаются и растягиваются стенки пузыря, сжимаются и расслабляются кольца
Акт движения рыб осуществляется за счет активной работы плавников. Плавники рыб бывают: парные-грудные, брюшные и непарные-спинной, анальный, хвостовой, спинной плавник может быть один (у карповых), два (у окуневых) и три (у тресковых). Хвостовой плавник принимает некоторое участие в поступательном движении, парализуя тормозящее движение конца тела и ослабляя обратные токи. По характеру действия хвосты рыб принято разделять на: 1)изобатический, где верхняя и нижняя лопасти равновелики: подобный тип хвоста имеется у скумбрии, тунца и многих других: 2)эпибатический – у которого верхняя лопасть развита лучше, чем нижняя: этот хвост облегчает движение вверх подобного рода хвост характерен для акул и осетровых: 3) гипобатический, когда нижняя лопасть хвоста развита больше верхней и способствует движению вниз: такой хвост имеется у летучей рыбы леща и некоторых других рыб. (Рис.66)
Жировой плавник без костных лучей представляет мягкий кожный вырост на задней части спины (лососевые). Плавники обеспечивают равновесие тела рыб и движения ее в разных направлениях, хвостовой плавник создает движущую силу и исполняет роль руля, обеспечивая маневренность рыбы при поворотах. Спинной и анальный плавники поддерживают нормальное положение тела рыбы, т.е. выполняют роль киля. Парные плавники поддерживают равновесие и являются рулями поворотов и глубин. Основную функцию рулей глубины выполняют у рыб грудные, а также брюшные плавники. С их помощью осуществляется отчасти и поворачивание рыбы в горизонтальной плоскости. Роль непарных плавников сводится к содействию поворотам рыбы вверх и вниз и к роли килей.
На брюшной стороне туловища, каудальное место прикрепления брюшных плавников имеется анальное отверстие, а сразу за ним – мочеполовой сосочек; у части рыб он представляет собой углубление с двумя отдельными отверстиями; мочевым (заднее) и половым. Место расположения заднепроходного, полового и мочевого отверстий за границей тазового и хвостового позвонка.
Плечевой пояс имеет вид хрящевого полукольца, лежащего в мускулатуре стенок тела позади жаберного отдела. Часть пояса называется лопаточным отделом, вентральнее – коракойдным. В основании скелета свободной конечности расположены 3 уплощенные базальные хрящи. Дистальнее базальных хрящей расположены в 3 ряда палочковидные радиальные хрящи. Остальная часть лопасти свободного плавника поддерживается многочисленными тонкими эластиновыми нитями – элементами вторичного кожного скелета.
Тазовый пояс – представлен хрящевой пластинкой, лежащей в толще брюшной мускулатуры перед клоакальной щелью. В брюшных плавниках имеется только один базальный элемент. Он сильно удлинен, и к нему прикрепляется один ряд радиальных хрящей. Остальная часть свободного плавника поддерживается эластиновыми нитями. У самцов удлиненный базальный элемент продолжается за пределы лопасти плавника как скелетная основа копулятивного выроста.
10.1.1 Непарные плавники. Представлены хвостовым анальным и спинным плавниками. У ключей акулы анального плавника нет. Хвостовой плавник акул гетероцеркальный, его верхняя лопасть значительно длиннее нижней, осевой скелет – позвоночник заходит только в верхнюю лопасть.
Скелетную основу хвостового плавника образуют удлиненные верхние и нижние дуги позвонков. Большая часть лопасти хвостового плавника поддерживается эластиновыми нитями. В основании скелета спинных и анальных плавников лежат радиальные хрящи, которые погружены в толщу мускулатуры. Иногда они сливаются в более крупные образования. Свободная лопасть плавника поддерживается эластиновыми нитями. У колючей акулы перед каждым спинным плавником расположен роговой шип, который, как и плакоидные чешуи, представляет собой элемент кожного скелета.
10.1.2 Жаберные крышки.Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой, и объединяющимися снизу при помощи непарных костных элементов. Пятая, самая задняя жаберная дуга, имеет лишь один парный (нижний) отдел.
Жаберные крышки состоят из четырех вторичных (покровных) костей: крышки (opezculum) и межкрышки (intezopezculum). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующей подвеске и к квадратной кости.
10.1.3 Парные конечности и их пояса.Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс. Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей – лопатка (scapula) расположена в области причленение свободной конечности. Сразу под ней находится коракоид (coracoideum). Эти два элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (coracoideum), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (supracleithum). Клейтрум в свою очередь соединяется с заднетеменной (posttemporale) костью. Направленные вперед нижние концы правого и левого клейтрумов соединяются друг с другом. Позади клейтрума неподалеку от лопатки и коракоида расположена небольшая заднеключичная кость (postcleithrum).
Все названные кости парные, они составляют вторичный плечевой пояс. Правая и левая заднетеменные кости причленяются к осевому черепу, что обеспечивает более прочную фиксацию пояса, и таким ообразом усиливает его опорную функцию.
Рис. 66. Скелет парных плавников и их поясов:
А – хрящевая рыба; Б – костистая рыба; 1 – грудной плавник с плечевым поясом;11 – брюшной плавник с тазовым поясом; 1 – лопаточный отдел; 2 – коракоидный отдел; 3 – базалии; 4 – радиалии; 5 – плавниковые лучи; 6 – птеригоподии; 7 –лопатка; 8 – коракоид; 9 – клейт-рум; 10 – задняя клейтрум; 11 – надклейтрум; 12 – задневисочная кость; 13 – тазовая кость
Грудной плавник в своем основании имеет один ряд мелких косточек – радиалий (radialia), отходящих от лопатки (частично и от коракоида). Вся свободная лопасть плавника состоит из членистых кожных лучей (lepidotrichia). Особенность скелета грудных плавников костистых рыб, по сравнению с хрящевыми, заключаются в редукции базалий. Подвижность грудных плавников увеличивается потому, что мышцы прикрепляются к расширенным основаниям кожных лучей, подвижно сочленяющихся с радиалами.
Тазовый пояс представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связан с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников лопасть плавника поддерживается кожными костными лучами (lepidotrichia), расширенные основания, которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников связано с их ограниченными функциями.
10.1.4 Непарные конечности.Непарные конечности представлены спинными, подхвостовыми (анальным) и хвостовым плавниками. Анальные и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птиригофоры и наружные плавниковые лучи – лепидотрихии.
Хвостовой плавник, имеет влияние равнолопастное строение, однако при рассмотрении его внутреннего скелета видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку – уростиль (urostal), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается расширенными, довольно широкими нижними дугами позвонков гипуралиями. Такой тип строения хвостового плавника, носит название галоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами – лепидотрихиями.
Из многочисленных способов движения известных среди животных, рыбам свойственны три: плавания, ползания и полет.
В зависимости от выполняемой функции, плавники делят на две группы. Первую группу составляют приспособления, непосредственно направленные на создание локомоторной силы, т.е. морфологические особенности, которые определяются отношением двигателя рыбы. Вторая группа- приспособления, которые движущаяся рыба встречает со стороны определяющей ее виды.
10.2 Функция локомоции и строение хвостового плавника.Функции хвостового плавника многообразны.
1. Хвостовой плавник участвует в общей локомоторной работе туловища, создавая силу, которая толкает рыбу вперед;
2. Действуя, как пассивная несущая полость, итероцеркальный хвостовой плавник создает некоторую подъемную силу, поддерживающую заднюю часть тела рыбы;
3. Хвостовой плавник является стабилизатором и отчасти вертикальным путем.
Хвостовой плавник у большинства рыб способен производить ряд сложных движений. Служит для изменения направления движения в вертикальной плоскости.
Первые три функции могут быть названы пассивными функциями хвостового плавника, так как при отправлении этих функций он действует просто как плоскость, более или менее метко прикрепленная на конце хвостового стебля. Последняя же функция является активной, так как при ее отправлении хвостовой плавник выполняет ряд сложных собственных движений и действует не просто как единая плоскость, а как сложная система, состоящая из отдельных элементов – лучей подвижно скрепленных с концом позвоночного столба. Особенности функции хвостового плавника у различных рыб определяют его форму, в отношении которой должны быть отличены в первую очередь два случая; хвостовой плавник, не разделенный на две лопасти, и хвостовой плавник, раздельный на две лопасти.
Плавание свойственно всем без исключения рыбам. Другие два способа характерны только для немногих видов, причем всегда имеют второстепенное значение. Наиболее общий и типичный для рыб способ движения плавания охватывает собой целый ряд модификации, которые могут быть разделены на две группы:
Первая группа заключает в себе способы плавания, в основе которых лежат волнообразные латеральные изгибания туловища.
Вторая группа – все остальные способы плавания, основу которых составляют те или иные движения плавников по характеру совершаемых плавательных движении делятся на 2 группы. Первая группа плавания основывается на движении плавников неундулирующего типа; преимущественно работа с грудными плавниками, при которых плавник отводится от тела в горизонтальном положении, затем ставится вертикально и приводится к телу, создавая в этот момент определенную локомоторную силу. Способ плавания Chronus chronus (L) u Crenilohrustina (L).Неундулирующие движения плавников являются второстепенным способом движения, при медленном плавании. Основным способом движения оно становится только у некоторых прибрежных медленно плавающих видов. Неундулирующие движения плавников используются в ряде случаев и для передвижения по грунту, как у Gohius иногда и для закапывания в грунт.
Другая группа имеет в своей основе ундулирующие движения плавников спинного и анального. Этот способ плавания встречаются у скатов. Громадному большинству рыб свойственно плавание при помощи латеральных изгибании, этот способ движения является для рыб закономерным. В основе локомоции при помощи ундулирующих движении плавников, лежат волнообразные изгибания пластинки плавника, обусловленные последовательности поперечности отношениями лучей.
Поступательное движение рыб в разных случаях обеспечивается ундуляцией грудных, спинного или анального плавников или же той или иной комбинацией действии этих плавников. У скатов единственным органом локомоции являются чрезвычайно расширенные грудные плавники. Большая поверхность этих плавников и значительная дорзовентральная подвижность их лучей превращает их в прекрасный ундулирующий аппарат. Волны ундуляции приходят по плавникам спереди назад, создавая движущую силу, направленную вперед. Дублирующими мембранами, сообщающими рыбе поступательные движения, являются спинной и анальные плавники. В этом случае совокупное действие этих плавников создает силу, движущую рыбу вперед, причем волны ундуляции на плавниках направлены параллельно линиям основания плавника. Благодаря волнообразным изгибаниям пластинки плавника, возникают силы, толкающие рыбу вперед.
10.2.1 Функции плавников. В теле рыбы выделяются четыре постоянные зоны, для которых специфичны определенные функции плавников. Эти зоны можно назвать функционально специфическими, так как для каждой из них специфична определенная функция или ряд функций. I — зона передних рулей, и несущих плоскостей, II — зона килей, III-зона стабилизаторов, и IV- зона задних рулей и локомоторных органов. Функции зоны I выполняются парными плавниками брюшных плавников или только грудными.
Функции зоны II выполняются той частью спинного плавника, которая расположена спереди от центра тяжести, а также брюшными плавниками и отчасти анальным, если эти плавники или их передние отделы расположены спереди от центра тяжести. Функции зоны II выполняются морфологическими обособленными отделами спинного и анального плавников, которые расположены сразу за центром тяжести.
Функции зоны III несут самые задние отделы спинного и анального плавников (если они приближены к заднему концу тела) и хвостовой плавник.
Указанное расположение на теле функционально специфических зон характерны для всех рыб вписывающих при помощи волнообразных изгибании тела.
10.3 Плавание рыб. Плавание рыб происходит благодаря работе поперечнополосатой мускулатуры, которая приводит в движение движительные плавники. Рыбы передвигаются в толще воды, создавая упор, т. е. отталкиваясь от среды, обладающей некоторой плотностью, вязкостью и податливостью. Проще всего понять явление упора, рассматривая гребное плавание при помощи парных грудных плавников.
Грудной плавник, закрепленный своим основанием на плечевом поясе, под действием мускулатуры совершает движения, подобные вращению калитки вокруг петель. При движении назад, во время гребка, плавник максимально расправлен и создает максимальный упор. При движении плавника вперед, при подготовке гребка, лучи сведены и создают минимальный тормозящий упор. Гребным плаванием пользуются щуки, зеленухи, колюшки, бычки, сростночелюстные и многие другие рыбы.
Одиночный цикл движения плавника может быть подразделен на движение вперед (подготовка гребка), движение назад (гребок) и период, когда плавник прижат к телу (рефрактерная фаза). Скорость плавания рыбы зависит от скорости движения плавника — скорость движения внешнего края плавника всегда больше скорости поступательного движения рыбы, т. е. плавник является двигателем. Угловая амплитуда движения плавника может достигать 900. Скорость растет при уменьшении доли времени рефрактерной фазы и при росте доли времени активных фаз — подготовки и гребка, которые занимают приблизительно одинаковую часть времени. Скорость плавания находится в прямой зависимости от частоты гребков, которая может достигать 20 Гц, но обычно не превышает 5 Гц.
Движение грудных плавников у большинства рыб поочередное (типа «кроль»), но у некоторых видов, например у карпа,— одновременное (типа «брасс»). Скорость плавания при работе одних только плавников небольшая, расход энергии тоже, по-видимому, небольшой, так как в работу вовлекаются небольшие массы мускулатуры плечевого пояса. Коэффициент полезного действия мускулатуры при таком типе плавания довольно высок.
При плавниковом плавании туловищно-хвостовая мускулатура не бездействует, она в какой-то степени напряжена для поддержания обтекаемой «позы». В противном случае тело начинает пассивно колебаться в набегающем потоке, как водоросль в потоке, или как флаг на ветру (так называемый флаттер), и это вызывает торможение.
Ограниченные массы мускулатуры вовлекаются в работу при плавании с помощью непарных плавников, например спинного, анального, когда эффективный упор создают поперечные складки плавников. Наибольшие скорости достигаются рыбами при периодическом волнообразном изгибании всего тела (ундуляции). Движителем в этом случае служит почти вся поверхность тела, за исключением негибкой головы.
При движении рыб активно сокращается почти вся туловищно-хвостовая мускулатура, составляющая приблизительно половину массы тела. Упор при ундуляционном плавании создается искривлением тела и движением локомоторной волны от головы к хвосту. За один цикл движения рыба могла бы продвинуться на длину тела до теоретического финиша, но в результате наличия КПД движителя 0,70 возникает пробуксовка и она проплывает расстояние до фактического финиша.
Н. В. Кокшайский охарактеризовал плавание как явление, при котором организм образует некоторую систему подвижных элементов, отталкивающих среду и одновременно отталкивающихся от нее. Длина локомоторной волны (расстояние между соседними гребнями) соизмерима с длиной рыбы.

Рис.67. Схематическое изображение плавания рыбы. Стрелки изображают направление приложения упора.
Скорость движения локомоторной волны по телу рыбы равна произведению длины волны на частоту взмахов хвоста, численно она также близка произведению длины рыбы на частоту ундуляции:
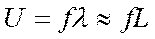
где U — скорость локомоторной полны; f—частота ундуляции; 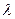 — длина локомоторной волны; L — длина рыбы. Вода представляет собой податливую среду, поэтому в ней имеет место некоторая «пробуксовка» движителей. Скорость плавания рыбы поэтому всегда меньше скорости локомоторной волны:
— длина локомоторной волны; L — длина рыбы. Вода представляет собой податливую среду, поэтому в ней имеет место некоторая «пробуксовка» движителей. Скорость плавания рыбы поэтому всегда меньше скорости локомоторной волны:
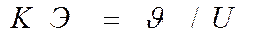
где КЭ — коэффициент эффективности движителя;  — скорость плавания рыбы.
— скорость плавания рыбы.
Максимальная скорость плавания рыбы является функцией длины тела и максимальной частоты ундуляции, т. е. максимальной частоты сокращенийтуловищной мускулатуры. Максимальная частота ударов хвоста при плавании некоторых достигает 30 Гц.(Таблица 20).
Максимальные частоты ундуляции и соответствующие им максимальные, так называемые спринтерские скорости могут поддерживаться в течение нескольких секунд. Далее наступает утомление и скорость резко падает, это связано со строением рыбы: с большим числом позвонков могут сильнее изгибать тело, чем рыбы с малым числом позвонков. Число позвонков у рыб колеблется поразному: так у рыбы -луны от 16 и у ремень рыбы до 400. Рыбы с мелкой чешуей также могут изгибать свое тело в большей степени, чем крупночешуйные.
Таблица 21 — Расчетные значения скорости плавания рыб (см./с) как функция частоты ундуляции (Гц) и длины рыбы (см) для области чисел Рейнольдса менее 105 (по Яржомбек, 1975)
| Длина рыбы | Частота ундуляции | |||||||||
| 0,1 1,2 5,7 | 0,7 3,4 | 0,2 2,4 8,4 | 0,7 4,5 | 1,3 6,8 | 2,1 9,2 | 4,8 | 6,9 | 9,6 |
Скорость движения рыб связана и с биологическим состоянием рыбы, в частности зрелостью гонад, температурой воды может меняться и от того, движется рыба в стае или в одиночку. Наибольших скоростей достигают некоторые акулы, меч- рыбы, тунцы. Голубая акула перемещается со скоростью около — 10 м./сек, тунец со скоростью 20м/ сек, лосось 5м/сек. Абсолютная скорость движения рыбы зависит от ее размеров. Крупные рыбы обладают большей скоростью, чем более мелкие. Эта зависимость выражается следующей формулой:
V = 1 [ L (3f-4)]
Где V— скорость в см./ сек.
L — длина рыбы в см.
f — число взмахов хвоста в секунду
Поэтому для сравнения скорости движения разноразмерных рыб используется обычно коэффициент скорости представляющий собой частное деление абсолютной скорости движения рыбы на корень квадратный ее длины (V).
Рыбы хорошие пловцы, в текучей воде по форме тела отличаются от хороших пловцов в стоячей воде, в частности у первых хвостовой стебель обычно значительно выше и короче, чем у вторых. В качестве примера можно сравнить форму хвостового стебля форели, приспособленной к жизни в воде с быстрым течением, и скумбрии- обитателя медленно двигающихся и стоячих морских вод.
Быстро плавая, преодолевая быстрины и перекаты, рыбы утомляются. Они не могут длительно плавать без отдыха. При большом напряжении у рыб в крови накапливается молочная кислота, которая после отдыха исчезает. Молочная кислота влияет на растворимость газов кровью и на транспортную функцию гемоглобина. Рыбы, при прохождении рыбоходов, утомляются, преодолевая их, даже гибнут.
Благодаря специфическому строению мускулатур рыб, сокращение каждого миомера вызывает изгиб тела на довольно большой протяженности, т. е. создается рычаг приложения силы. Мышечные волокна в миомерах ориентированы таким образом, что одно волокно является как бы продолжением другого и такие составные нити косо расходятся от средней линии по направлению к хвосту и краям тела.
Светлая и темная мускулатура рыб во многом различна. При спокойном плавании рыбы в крейсерском режиме число сокращений волокон темной мускулатуры равно числу плавательных движений. Сокращения волокон белой мускулатуры носят нерегулярный характер.
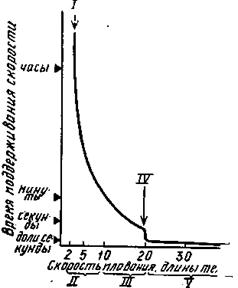 Существует некоторая неутомляющая частота мышечных сокращений, при которой времени между двигательными актами достаточно для полного восстановления работоспособности. При таких режимах движения происходят длительные миграции рыб. Неутомляющая средняя скорость в 5—10 раз меньше максимальной спринтерской скорости плавания. Проведенные эксперименты и наблюдения в естественных условиях показали, что лососи не утомляются при скорости плавания 1—2 длины тела в секунду, т. е. до 5 км/ч.
Существует некоторая неутомляющая частота мышечных сокращений, при которой времени между двигательными актами достаточно для полного восстановления работоспособности. При таких режимах движения происходят длительные миграции рыб. Неутомляющая средняя скорость в 5—10 раз меньше максимальной спринтерской скорости плавания. Проведенные эксперименты и наблюдения в естественных условиях показали, что лососи не утомляются при скорости плавания 1—2 длины тела в секунду, т. е. до 5 км/ч.
Рис.68. Скорости плавания быстроходных рыб: I- неутомляющая скорость;II- стайерская скорость; III- скорость средних дистанции; IV- спринтерская скорость; V- скорость прыжков с разбега
Тунцы — рекордсмены среди рыб; по скоростной выносливости могут длительное время двигаться со скоростью 3—4 длины тела в секунду, т. е. порядка 20 км/ч.
Между неутомляющими и спринтерскими скоростями имеется область, где время поддерживания скорости находится в обратной связи с заданной скоростью.
По аналогии со спортивной терминологией эту область можно разбить на «средние дистанции» — до времени движения несколько десятков минут и «стайерские дистанции». Стайерская скорость может поддерживаться несколько часов, но она все-таки не является неутомляющей, или крейсерской, скоростью (рис. 69).
Наилучшие среди рыб пловцы — тунцы, ставриды, лососи. Они плавают в 2-3 раза быстрее и выносливее, чем посредственные пловцы, такие, как осётровые, камбалы, бычки, угри.
Таблица 22 — Максимальная частота плавательных движений
§
| Вид рыбы | Длина, см. | Максимальная, известная частота, Гц | Вид рыбы | Длина, см. | Максимальная известная частота, Гц | |
| Тунец Вьюн Бычок Карп | Хамса Атерина Колюшка Гольян |
|
| |||
У крупных хищников и мигрантов абсолютные значения максимальной скорости могут достигать больших величин. Двухметровая рыба, работающая с частотой 30 Гц и движущаяся со скоростью около 20 длин тела в секунду, развивает скорость 40 м/с, или 144 км/ч. Одиночные мощные разгибы тела могут происходить в течение промежутков времени короче 1/30 сек. В этот момент может происходить еще более быстрое — до 40 длин тела в секунду, но очень кратковременное перемещение. По аналогии со спортом такие рывки можно назвать прыжками с разбега.
10.4 Полет рыб.Среди рыб есть представители, которые приспособились к своеобразному полету по воздуху, это не настоящий полет, а парение по типу планера. У этих рыб грудные плавники чрезвычайно сильно развиты и выполняют ту же функцию, что крылья самолета или планера. Основной двигатель, дающий начальную скорость при полете, хвост и в первую очередь его нижняя лопасть. Выскочив на поверхность воды, летучая рыба еще некоторое время скользит по водной поверхности, оставляя за собой кольцевые волны, расходящиеся в стороны. В то время, когда тело летучей рыбы находится в воздухе, а в воде остается только ее хвост, она все еще продолжает увеличивать скорость движения, нарастание которой прекращается только после полного отрыва тела от поверхности воды.
Держаться в воздухе летучая рыба может около 10-15 сек. поднимаясь в высоту до 4м. над поверхностью воды, пролетая при этом расстояние свыше 100 м. Дальше держаться в воздухе могут те рыбы, у которых увеличены как грудные, так и брюшные плавники.
В момент взлета грудные плавники рыбы широко раскрыты и расположены под углами к направлению движения, благодаря чему создают необходимую подъемную силу. Эта функция и обусловливает их очень сильное развитие.
Полет у летучих рыб выработался как защитное приспособление, позволяющее им ускользать от хищников — тунцов, корифен, меч — рыб. Полет рыб есть так же приспособление к перемещению с меньшей затратой энергии от одного скопления кормовых планктонных организмов к другому.
Среди харациновых рыб есть представители, приспособившиеся к активному машущему полету. Это мелкие рыбки до 9-10см длины, населяющие пресные воды южной Америки. Они могут выскакивать из воды и пролетать при помощи взмахов удленненных грудных плавников до 3-5 метров. Хотя у летающих харацинид размеры грудных плавников меньше, чем у летучих рыб данного семейства. Грудные мышцы, приводя в движение грудные плавники, хорошо развитые у харациновых рыб, приспособившихся к очень сильно развитым костям плечевого пояса, которые образуют некоторое подобие грудного киля птиц. У летающих рыб плавники достигают 25% веса тела, в то время как у нелетающих представителей близкого рода — только 0,7%.
Другим способам движения рыб можно отнести движения при помощи прикрепления к плавающим предметам паразитизма, комменсализма и симбиоза. Изменением направления двигателя, сохранением равновесия и торможением, морфологически касаются главным образом особенностей строения плавников. Обособляются плавники, выполняющие функции несущих плоскостей, рулей килей стабилизаторов и локомоторных органов, причем расположение всех этих плавников на теле строго подчинено генетическому коду.
10.5 Общая характеристика скелетной мускулатуры.
Движение—одно из проявлений жизни; у позвоночных животных оно осуществляется специальными органами—мускулами, построенными из поперечнополосатой мышечной ткани. Мускулы закрепляются на скелете на системе рычагов и в целом составляют скелетную мускулатуру.
Своей работой скелетная мускулатура обеспечивает: а) перемещение тела животного в пространстве (в поисках пищи, особи другого пола, в целях защиты); 6) изменение положения отдельных частей тела относительно друг друга; в) изменение объема полостей тела (при дыхании и пр.); г) изменение положения и направления складок кожи (движение рта и т. д.).Движения животного являются ответом на раздражения, полученные из внешней среды, мышечные движения — одно из внешних проявлений мозговой деятельности (И. М. Сеченов).
Мышечная система в значительной степени влияет на строение и развитие органов активного движения в целом, и в зависимости от степени и характера ее собственного развития происходят соответствующие изменения в строении скелета.
Наконец, мышечная система служит трансформатором химической энергии, поступающей в организм в виде пищи, в тепловую (до 70%) и в механическую работу (всего лишь около 30%).
10.5.1 Морфология и физиология мышц (мускула). Мускул, или мышца,—musculus—имеет соединительнотканный остов и главные функционирующие элементы, которыми служат поперечнополосатые мускульные волокна.
Соединительнотканному остову присущи общие для всех органов черты построения, т. е. снаружи мускул одет соединительнотканной оболочкой—наружным перимизием—perimysium extcrnum s. epimysmm — который иногда содержит небольшое количество жировой ткани. От этой оболочки проходят внутрь органа значительные по толщине перегородки, или; трабекулы (2), связывающие между собой крупные мускульные пучки. Перегородки, в свою очередь, отдают от себя ряд более тонких перегородок, которыми охватываются менее крупные мускульные пучки. Все эти внутренние переплеты, по которым ветвятся сосуды и нервы, получают название внутреннего перимизия—perimysium internum (3). Между мускульными волокнами самых мелких мускульных пучочков также пробегают очень нежные прослойки, называемые ондомизием—endomysium. Оболочка вместе со всеми указанными перегородками и составляет остов мускула, в который закономерно вплетены мускульные волокна. Все эти образования в совокупности и дают различной величины и формы мускульное брюшко.
Каждое мускульное брюшко имеет сухожильный добавок—сухожилие. Форма сухожилия зависит от формы мускульного брюшка. Если мускульное брюшко в сечении округлой формы, то и сухожилие как его добавок становится шнурообразным. В таких случаях его принято называть просто сухожилием- tendo (tenon). Если же мускул плоский, то он переходит в пластинчатое сухожилие-апоневроз.
Сухожильные добавки построены наподобие самого мускульного брюшка, но вместо пучков сократительных элементов здесь находятся
параллельные волокна плотной оформленной соединительной ткани. Эти волокна идут пучками и заложены в свой остов, который является продолжением остова брюшка, т. е. оболочка и трабекулы брюшка—наружный и внутренний перимизии—переходят в оболочку и трабекулы сухожилия— peritendmeum (peri-tenonium) extcrmim et peritendmeum internum. Волокна сухожилия крепко соединены с мускульными волокнами (а может быть, и переходят непосредственно друг в друга). Сухожилие становится очень плотным (компактным) и прочным шнуром благодаря наличию лишь крайне незначительной соединительнотканной прослойки между строго параллельно направленными коллагенными пучками волокон, скудно снабженными сосудами. Все это делает сухожилие мало утомляемой частью мускула. Кроме того, сухожилие по сравнению с мускульным брюшком обладает огромной сопротивляемостью на растяжение (Рис.70).
Сухожилия весьма крепко соединены с рычагами движения, проникая в виде шарпеевских волокон в толщу костного вещества, с такой конструкцией и свойствами сухожилие принимает на себя функцию передатчика силы сокращения мускульного брюшка на отдаленные, скелетные части, т. е. играет роль трансмиссионного приспособления.
Вступающие в мускул и выходящие из него сосуды распределятся в глубину по перекладинам остова. Артерии, подходя к мускульным волокнам, образуют густую сеть капилляров, петли которой растянуты в направлении длинной оси волокон.
Мускул снабжен двигательными и чувствительными нервными волокнами. Каждое отдельное волокно мускула управляется толькоему присущим самостоятельным двигательным волокном и вместе с ним представляет одну целую, очень маленькую рабочую единицу. Это единство устанавливается у зародыша с самых начальных стадий развития мускульных клеток (волокон).
Таким образом, комплекс объединенных мускульных волокон дает вместе с остовом и сухожилием орган-мускул, который, согласно сказанному, неразрывно связан со своими нервными волокнами, составляющими целый нерв данного мускула.
В процессе филогенеза при функциональной дифференцировке мускула последний может распадаться на новые мускульные единицы, и этому распадению обязательно следует и сопровождающий его нерв. При обратных преобразованиях, т. е. при слиянии в результате объединения работы нескольких мускулов в один сплошной орган, к нему будут подходить, и его будут иннервировать несколько нервов, и мускул станет по происхождению сложным. Если принять в соображение, что каждый мускул развивается из определенного мускульного сегмента (миотома или его вентрального отпрыска) и иннервируется нервом того же сегмента, то легко понять, что иннервация мускулатуры различными ветвями служит очень ценным критерием при определении происхождения и гомологии отдельных мускульных единиц.
Форма мускулов. Величина и форма мускулов чрезвычайно разнообразны и тесно связаны с взаиморасположением плечей рычагов, на которых укрепляются мускулы, и силой сокращения в том или другом месте организма.
Мускулы особенно рельефно обрисованы на свободных конечностях. По сравнению с мускулами туловища и головы они имеют в большинстве случаев удлиненную, в сечении округлую форму, напоминая или цилиндр, или конус, или веретено. На поясах конечностей эта рельефность формы и обособленности уже несколько стушевывается, а мускулы, простирающиеся между туловищем и плечевым поясом, характеризуются в большинстве пластинчатой формой, т. е. представляют так называемые широкие мускулы. Мускулы, располагающиеся вокруг отверстий, бывают кольцеобразной формы(Рис.71).
Внутреннее строение мускулов.Внутреннее строение мускульных брюшков бывает довольно разнообразно, наиболее простым видом строения следует считать брюшко, у которого направление волокон параллельно длинной оси мускула. Такого типа строения мускулы очень мясистые и называются по функции динамическими, а по строению простыми. Мускул тем сильнее, чем больше в нем мышечных волокон. Количество волокон практически сосчитать нельзя, поэтому силу мускула принято определять путем измерения его физиологического поперечника. В простых мускулах, как уже отмечалось, все пучки мышечных волокон параллельны длинной оси мускула; следовательно, в данном случае физиологический поперечник совпадает с физиологическим и анатомическим поперечником, который также перпендикулярен длинной оси мускула.
В зависимости от гистологического строения подъемная сила мускула может колебаться в пределах от 5 до 15 кг (в среднем 8—10 кг) на 1 см² физиологического поперечника. Расстояние, на которое мускул может проявить свою силу, зависит от длины мышечных волокон, которая не превышает 12—15 см. Так как мышечное волокно при своем сокращении укорачивается приблизительно на половину своей длины, то мускул, имеющий физиологический поперечник в 1 см2, способен поднять груз в 10 кг на высоту 6—7 см, т. е. механическая работа мускула в этих условиях будет равняться приблизительно 0,6 кг/м.
Мускулы описанного динамического типа, будучи маломощными, встречаются крайне редко. Чаще мышечные волокна имеют косое направление по отношению к длинникумускула, располагаясь при этом в нескольких вариациях.
1. Начальное и концевое сухожилиямогут покрывать мускульное брюшко с его противоположных поверхностей на большем или меньшем протяжении. Они обычно хорошо видны и за свой блестящий белый цвет называются мышечными (сухожильными) зеркалами. В отношении к длиннику мускула пучки мышечных волокон идут косо от одного сухожилия к другому или от кости (если мускул начинается мясисто) к концевому сухожилию. Такие мускулы называются одно-или полуперистыми— m. unipermatus. Вполне понятно: а) что при одинаковых объемах простого и перистого мускулов количество мышечных волокон в последнем будет больше, но длина соответственно меньше. Следовательно, мускул будет выигрыватьв силе, но проигрывать в расстоянии; б) что физиологический поперечник (с~с) в перистых мускулах не будет соответствовать анатомическому (в—в)— последний меньше.
2. Одно из сухожилий может расщепляться на две пластины, которые покрывают один конец мышечного брюшка, а в толщу последнего внедряется противоположное сухожилие. В таком случае мышечные волокна будут косо подходить к центральному сухожилию с двух противоположных поверхностей. Такой мускул по своему строению—д вуперисты й—m. bipennatus. В нем будет еще больше мышечных волокон, а длина, напротив, еще меньше.Двуперистый мускул еще больше выигрывает в силе за счет большого проигрыша в расстоянии. Наиболее сложное устройство приобретает мускул, когда расщепляет сухожилия; например, одно сухожилие на три пластины, и тут создается громадная подъемная сила, действующая, на очень коротком расстоянии.
Двухперистые и многоперистые мускулы называются статодинамическим и, так как они по преимуществу выполняют статическую функцию (например, при упоре животного). Вместе с тем такие мускулы (перистые) при соответствующих условиях могут давать также очень значительный размах. К таким условиям может быть отнесено использование более короткого плеча силы рычага, чем это требуется для динамического мускула. При этом такие мускулы дают очень сильный, хотя и короткий первоначальный импульс движению, который обычно совпадает с моментом наибольшей потребности в силе сокращения и способен по инерции увеличить размах плеча сопротивления.
Рис. 70. Типы построения мускульных брюшков
А — одноперистый тип;
Б — двуперистый тип;
С — многоперистый тип;
а- сухожилие;е-е- анатомический
и с- с- физиологический поперечник.
Механизм действия мускулов.Действие мускулов выражается их сокращением, причем химическая энергия мускульной тканью превращается в работу с выделением теплоты. При сокращении брюшко мускула укорачивается по длинной оси приблизительно наполовину, а в перистых мускулах очень мало, и утолщается в поперечнике. Это влечет за собой сближение пунктов прикрепления на рычагах, а следовательно, и самих рычагов (рис.70). Такое сближение достигается различными способами.
Во-первых, как это бывает чаще всего, один из пунктов прикрепления остается неподвижны м-punctum fixum-и в результате сокращения перемещается лишь та кость или часть тела, на которой находится подвижное прикрепление-punctum mobile.
Во-вторых, наблюдаются случаи, когда места прикрепления в функциональном отношении могут изменяться, т. е. неподвижное место при одной потребности передвижения может стать подвижным, а при другой потребности, наоборот, неподвижным.
Существуют и такие условия, при которых начало и конец мускула совсем отсутствуют, как, например,у мускулов, окружающих кольцом отверстия (ротовое) или полые органы (сфинктер прямой кишки у анального отверстия).
Закономерные действия мускулов или их групп в системе органов произвольного движения обусловливают самые разнообразные изменения в соотношении частей тела друг с другом и всего организма с внешней средой (перемещение по суше, в воде, в воздухе).
В частности, мускульная работа вызывает движения: 1) друг относительно друга частей позвоночного столба, 2) грудных и брюшных стенок, 3) нижней челюсти и подъязычной кости относительно дорзального отдела черепа и кожных складок головы 4) поступательные и хватательные конечности. Смотря по эффекту, получающемуся в результате мускульного сокращения, на конечностях принято различать следующие мускулы: а) сгибатели и разгибатели, б) приводящие и отводящие, в) вращатели наружу ивращатели внутрь и г) напрягатели. Эти определения вполне совпадают с теми видами движения, которые уже рассмотрены по отношению ккаждому типу суставов.
Каждая ось движения имеет лишь две мускульные группы, из которых одна действует на сустав в направлении, диаметрально противоположном другой. Отсюда становятся понятными употребляемые по отношению к мускулам названия антагонистов—antagonist—и синергистов, или сотоварищей,—synergetes s. socii. Так, например, разгибатели какого-либо сустава увеличивают его угол, а сгибатели уменьшают, следовательно, разгибатели и сгибатели будут друг другу антагонистами; мускулы, действующие на сустав только в одном направлении, например, сгибая его, будут друг другу синергистами.
Мускул обладает очень совершенной упругостью, вследствие чего он даже в нерабочие моменты находится в слегка растянутом, т. е. напряженном, состоянии (в состоянии мышечного тонуса) и вполне гарантирован от удлинения после своего растяжения действием антагонистов. Эластичность брюшка предохраняет также от порывистых, дергающих движений рычагов при начале сокращения. Этим создается плавная работа на рычагах и устраняется нецелесообразная трата сил на предварительное натяжение самого мускульного брюшка перед сокращением.
Фасции.Фасции—fascia—представляют собой оболочки или пластины, построенные из плотной соединительной ткани. Они формируют для мускулатурыв целом, а также для некоторых других органов дополнительный к скелету фиброзный сложный пластинчатый остов. В его пластины заключены как отдельные мускулы, так и мускульные группы и, наконец, вся мускулатура и сосудисто-нервные пучки.
Общее распределение этого фиброзного остова таково. Все тело животного под кожным покровом одето, как чехлом, подкожной, или поверхностной фасцией—fascia subcutanea s. superficialis. Эта фасция имеет отношение главным образом к подкожным мускулам туловища.
Под поверхностной фасцией лежит второй сложный чехол— глубокая фасция—fascia profunda, —которая уже непосредственно прилегает к поверхностным скелетным мускульным группам, одевая последние в оболочку.
Мускулатуры туловища и хвоста.Среди хордовых животных мускулатура туловища наиболее примитивно построена у ланцетника. За некоторым изъятием она представляет в общем два сильноразвитых, но не дифференцированных мускула — правый и левый. Каждый из них имеет миомерию, т. е. расчленен на десятки (до 60) сегментов поперечными соединительнотканными перегородками. Этасегментация распространяется и на область жаберного аппарата, располагаясь дорзально от него. Короткие пучки мускульных волокон между поперечными перегородками всюду идут исключительно в продольном направлении.
У стоящих выше по зоологической лестнице рыб, одновременно с резким обособлением головы и плавникообразных конечностей мускулатура тела распадается на мускулы головы, туловища, хвоста и конечностей. В мускулатуре туловища и хвоста с такой же ясной миомерией, как и у ланцетника, на каждой симметричной половине появляется уже по продольной боковой перегородке, отъединяющей дорзальную мускулатуру от вентральной. Дорзальныймускул—m. dorsalis,—иннервируемый дорзальными ветвями спинномозговых нервов, в общем напоминает простые отношения у ланцетника, причем краниальный отдел его заканчивается на черепе и дорзальной части плечевого пояса. Вентральныймускул—m. ventralis,— обслуживаемыйвентральными ветвями спинномозговых нервов, у некоторых рыб начинает дифференцироваться. Эта дифференциация касается только области туловища, так как хвостовой отдел сохраняет простоту миомерии и продольное направление мускульных пучков.
Изменения на туловище выражаются в выделении у средней сагиттальной линии живота парного прямого мускула—m.rectus—с ясной миомерией; на боковых сторонах (как у ганоидных, так и у костистых рыб) мускульная масса, сохраняя миомерию, расчленяется на два косых пласта—поверхностный и глубокий. Поверхностный пласт представляет собой косой наружныймускул—m. obliquus externus; его пучки идут спереди и сверху назад и вниз. Глубокий пласт в виде косого внутреннего мускула—m. obliquus interims,— пучки которого направлены сзади и сверху вперед и вниз, перекрещивает первый. Сегментальное расчленение остается все же ясно выраженным и по всей вентральной мускулатуре.
У головы рыб из вентральной мускулатуры выделяется висцеральная мускулатура. Она направляется от коракоидной кости к нижней челюсти, подъязычной дуге и жаберным дугам. К челюсти у средней сагиттальной линии идет m. coracomandibularis, рядом с ним лежит идущий к подъязычной кости
Мускулатура рыб.Мускулатура — это специализированная сократительная ткань, которая подразделяется на два основных типа: поперечнополосатую произвольную и гладкую непроизвольную мускулатуру.
Мясо свежей пресноводной рыбы большинства пород содержит влаги—70—80%, белков—17—19%, солей— 1—1,3%, жира—от 0,5 до 10%, у некоторых рыб несколько больше. Содержание жира зависит от возраста рыбы, ее питания и время вылова, рыба осенью имеет больший процент жира, чем весной. У рыб лососевых пород жирность доходит до 20—24%.
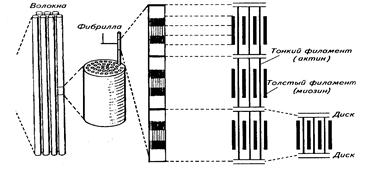 Гладкая непроизвольная мускулатура у рыб работает независимо от волевых актов, хотя тоже иннервирована и подчиняется действию нервных и гормональных стимулов. Гладкая мускулатура способна к спонтанным ритмическим сокращениям даже вне организма.
Гладкая непроизвольная мускулатура у рыб работает независимо от волевых актов, хотя тоже иннервирована и подчиняется действию нервных и гормональных стимулов. Гладкая мускулатура способна к спонтанным ритмическим сокращениям даже вне организма.
Рис.71.Продольное сечение быстрой скелетной мышцы
Например, полный цикл периодического движения желудка и кишечника рыб при комнатной температурре занимает несколько минут. Раздражение ветви, блуждающего нерва, иннервирующей пищеварительный
тракт, вызывает задержку сократительной деятельности. Характеристика деятельности плавательного, мочевого, желчного пузырей рыбы не имеет принципиальных отличий от работы гладкой мускулатуры кишечника. Для работы этих органов не требуется мгновенной реакции и мощного действия. Сокращение гладкой мускулатуры стимулируется многократными нервными и электрическими импульсами. Быстрое растяжение гладких мыщечных волокон под действием внешних сил, например растяжение желудка проглоченной пищей, вызывает ответное сокращение. В стенках полых органов (желудка, кишечника, пузырей) имеются продольные и кольцевые гладкие мышечные волокна. Гладкая мускулатура имеется также в селезенке, радужке глаза и в аппарате движения хрусталика глаза, в стенках кровеносных сосудов и жабрах.
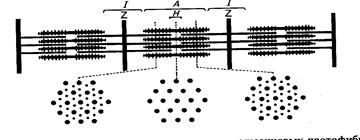 Рис.72.Схема строения попе-
Рис.72.Схема строения попе-
речнополосатой мышцы с перекрывающими друг друга рядами тонких и толстых нитей, с поперечными мостиками и Z пластинками в середине 1-дисков.Вверху- продольный разрез, внизу-поперечный, через разные участки саркомера.
СТРОЕНИЕ И РАБОТА ПОПЕРЕЧНОПОЛОСАТОЙ МУСКУЛАТУРЫПоперечно-полосатая произвольная мускулатура, или туловищно-хвостовая, является основным двигателем при плавании рыб. Деятельность ее в значительной мере определяется волевыми актами, хотя при плавании большую роль играет автоматизм.
Туловищно-хвостовая мускулатура рыб состоит из отдельных сегментов (миомеров), число которых равно числу позвонков. У рыб вытянутой формы число миомеров около 100 и больше (угорь), у рыб обыкновенной формы (карп, окунь,сельдь, лосось, камбала) примерно 50, у коротких рыб (бычки спинороги, ставрида) около 25 миомеров. Сегментарное строение характерно как для красной, или темной, латеральной мускулатуры, так и для белой, или светлой, составляющей большую часть всей мышечной массы. Несмотря на сегментарное строение, красная мускулатура выступает как единое целое. Все ее мышечные волокна направлены вдоль оси тела. Сокращение этих волокон вызывает латеральные изгибы тела. Белая мускулатура может быть разделена на спинную — выше средней линии тела — и боковую — ниже средней линии тела.
Соединительнотканные элементы, отделяющие спинную мускулатуру от боковой в один миомер, называются миосептами. К ним присоединяются мышечные волокна, с помощью которых сокращение мускулатуры передается костному скелету и вызывает локомоторные волнообразные движения рыбы.
Миомеры имеют сложную геометрическую форму, поэтому поперечный разрез тела рыбы рассекает несколько миомеров, представляющих собой как бы серию вложенных один в другой конусов, толщина которых приблизительно равна длине позвонка. У тунцов и акул таких слоев бывает 17—18. Непосредственно после импульса наблюдается короткий латентный (скрытный) период длительностью в несколько милли секунд. Затем происходят быстрое сокращение и более медленное расслабление. Время, требующееся для полного одиночного цикла, колеблется от 30 до 100 милли секунд.
Рис.74. а – миксина; б – акула; в — судак; 1,2- миомеры; 3-миосепты; 4-спинная мускулатура; 5 — латеральная мускулатура; 6-мышечные конусы направленные вперед; 7 — мышечные конусы, направленные назад; 7- поверхностная темная мускулатура; 9-оперкульарно -челюстная группа мышц; 10-группа мышц, сводящих и разводящих лучи хвостового плавника.
Если подавать на мышечное волокно стимулирующие импульсы до конца полного расслабления, то наблюдается продолжительное — изометрическое сокращение, или тетанус. Тетанус бывает частичный и полный. Изометрические напряжения мускулатуры несвойственны рыбам в обычных условиях существования. Они наблюдаются при экспериментальных работах и при электролове. Полный тетанус светлых волокон у ската наступает при стимуляции с частотой 5— 10 Гц, а максимальное напряжение развивается в изометрическом сокращении при частоте 20 Гц. Тетанус быстрых волокон у трески наблюдается при стимуляции с частотой 35—45 Гц, а максимальное напряжение — при частоте стимуляции 250— 400 Гц. По-видимому, гладкий тетанус мышечных волокон наступает при частоте стимуляции, приблизительно в 2—3 раза превышающей максимальную частоту произвольных плавательных движений.
При электрическом раздражении туловищной мускулатуры целой рыбы можно наблюдать тетанус, т. е. судорожное напряжение мускулатуры, но он маскируется множеством нерегулярных подрагиваний. Это явление можно наблюдать, если приложить стимулирующие электроды к боковой поверхности карпа и подавать электрические импульсы с частотой 50 Гц и амплитудой 5 В.
Естественные конвульсивные движения наблюдаются во время нереста у многих видов рыб. Изгибаются — и дрожат во время «брачных игр» лососевые, карповые и некоторые другие рыбы. Титаническое сокращение тела наблюдается при икрометании и выделении молок.
Коэффициент полезного действия плавательной мускулатуры рыб, по всей видимости, не превышает 25%. Коэффициент полезного действия двигательной системы сперматозоидов, как показывают энергетические расчеты, имеет близкую величину — порядка 20%. Это показывает, что сократительные элементы в организме имеют одинаковую физическую и химическую природу.
Мышечная система и электрические органы.Мышечную систему рыб, как и других позвоночных, разделяют на мышечную систему тела (соматическую) и внутренних органов (висцеральную). В первой выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру. Мышечная система взаимосвязана со скелетом (опора при сокращении) и нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определенным нервом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышц, которая в отличие от мышц млекопитающих невелика.
У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. Она обеспечивает плавание рыбы. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца — m. lateralis magnus) (рис. ). Продольной соединительнотканной прослойкой эта мышца делится на спинную (верхнюю) и брюшную (нижнюю) части.
Боковые мышцы разделены миосептами на миомеры, число которых соответствует количеству позвонков. Наиболее отчетливо миомеры видны у личинок рыб, пока их тела прозрачны. Мышцы правой и левой сторон, поочередно сокращаясь, сгибают хвостовой отдел тела и изменяют положение хвостового плавника, благодаря чему тело двигается вперед.
Над большой боковой мышцей вдоль тела между плечевым поясом и хвостом у осетровых и костистых лежит прямая боковая поверхностная мышца (m. rectus lateralis, m. lateralis superficialis). У лососевых в ней откладывается много жира. По нижней стороне тела тянется прямая брюшная мышца (m. rectus abdominalis); у некоторых рыб, например, угрей, ее нет. Между ней и прямой боковой поверхностной мышцей располагаются косые мышцы (m. obliguus).
Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура), плавники имеют свою мускулатуру. Наибольшее скопление мускулов определяет и расположение центра тяжести тела: у большинства рыб он находится в спинной части.
Деятельность туловищных мышц регулируется спинным мозгом и мозжечком, а висцеральная мускулатура иннервируется периферической нервной системой, возбуждаемой непроизвольно.
Различают поперечнополосатые (действующие в значительной степени произвольно) и гладкие мышцы (которые действуют независимо от воли животного). К поперечно — полосатым относятся скелетные мышцы тела (туловищные) и мышцы сердца. Туловищные мышцы могут быстро и сильно сокращаться, однако скоро утомляются. Особенностью строения сердечных мышц является не параллельное расположение обособленных волокон, а разветвление их кончиков и переход из одного пучка в другой, что обусловливает непрерывную работу этого органа.
Гладкие мышцы также состоят из волокон, но гораздо более коротких и не обнаруживающих поперечной исчерченности. Это мышцы внутренних органов и стенок кровеносных сосудов, имеющие периферическую (симпатическую) иннервацию. Поперечнополосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина — белка, легко связывающего кислород. Миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии. Красные и белые волокна различны по целому ряду морфофизиологических характеристик: цвету, форме, механическим и биохимическим свойствам (интенсивность дыхания, содержание гликогена и т. д.). Волокна красной мышцы (m. lateralis superficialis) — узкие, тонкие, интенсивно кровоснабжаемые, расположенные поверхностно (у большинства видов под кожей, вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина; в них обнаружены скопления жира и гликогена.
Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых. В мышце сердца (красной) мало гликогена и много ферментов аэробного обмена (окислительный обмен). Она характеризуется умеренной скоростью сокращений и утомляется медленнее, чем белые мышцы. В широких, более толстых, светлых белых волокнах m. lateralis magnus миоглобина мало, меньше в них гликогена и дыхательных ферментов.
Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко. Красные мышцы постоянно деятельны. Они обеспечивают длительную и непрерывную работу органов, поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца. При быстром движении, бросках активны белые мышцы, а при медленном — красные. Поэтому наличие красных или белых волокон (мышц) зависит от подвижности рыб: «спринтеры» обладают почти исключительно белыми мышцами. У рыб, которым свойственны продолжительные миграции, кроме красных Боковых мышц имеются добавочные красные волокна в белых мышцах.
Основную массу мышечной ткани у рыб составляют белые мышцы. Например, у жереха, плотвы, чехони на их долю приходится 96,3; 95,2 и 94,9% соответственно. Белые и красные мышцы различаются по химическому составу. В красных мышцах содержится больше жира, тогда как в белых мышцах больше влаги и белка. Толщина (диаметр) мышечного волокна изменяется в зависимости от вида рыб, их возраста, величины, образа жизни, а у прудовых рыб — от условий содержания. Например, у карпа, выращенного на естественной пище, диаметр мышечного волокна составляет (мкм): у мальков — 5-19, сеголетков — 14-41, двухлетков — 25-50.
Туловищная мускулатура образует основную долю мяса рыбы. Выход мяса в процентах общей массы тела (мясистость) неодинаков у разных видов, а у особей одного вида различается в зависимости от пола, условий содержания и др. Мясо рыб усваивается быстрее, чем мясо теплокровных животных. Оно чаще бесцветно (судак) имеет оттенки (оранжевый — у лососевых, желтоватый у осетровых и др.) в зависимости от наличия различных жиров и каротиноидов. Основную массу белков мышц рыб составляют альбумины и глобулины (85%), всего же у разных рыб выделяют 4 — 7 фракций белков.
Химический состав мяса (вода, жиры, белки, минеральные вещества) различен не только у разных видов, но и в разных частях тела. У рыб одного вида количество и химический состав мяса зависят от условий питания и физиологического состояния рыбы. В нерестовый период, особенно, у проходных рыб, расходуются резервные вещества, наблюдается истощение и, как следствие, уменьшается количество жира и ухудшается качество мяса.
У кеты, например, во время подхода к нерестилищам относительная масса костей увеличивается в 1,5 раза, кожи — в 2,5 раза. Мышцы оводняются — содержание сухого вещества снижается более чем в два раза; из мышц практически исчезают жир и азотистые вещества — рыба теряет до 98,4% жира и 57% белка. Особенности окружающей среды (в первую очередь пищи и воды) могут сильно изменять пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом. Качество мяса зависит и от диаметра мышечного волокна, а также количества жира в мышцах. В значительной мере оно определяется соотношением массы мышечной и соединительной тканей, по которому можно судить о содержании в мышцах полноценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки). Это соотношение изменяется в зависимости от физиологического состояния рыбы и факторов внешней среды.
В мышечных белках костистых рыб на белки приходится: саркоплазмы 20 — 30%, миофибрилл — 60 — 70, стромы около 2%. Все многообразие движений тела обеспечивает работа мышечной системы. Она, главным образом, обеспечивает и выделение тепла и электричества в организме рыбы. Электрический ток образуется при проведении нервного импульса по нерву, при сокращении миофибрилл, раздражении светочувствительных клеток, механохеморецепторов и др.
Электрические органы. Своеобразно измененными мышцами являются электрические органы. Эти органы развиваются из зачатков поперечно-полосатой мускулатуры и расположены по бокам тела рыб. Они состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки (электроциты), переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя — положительно. Разряды происходят под действием импульсов продолговатого мозга. Вследствие разрядов вода разлагается на водород и кислород, поэтому, например, в заморных водоемах тропиков около электрических рыб скапливаются мелкие обитатели — моллюски, рачки, привлеченные более благоприятными условиями дыхания.
Электрические органы могут располагаться в разных частях тела: например, у ската, морской лисицы — на хвосте, у электрического сома — на боках. Генерируя электрический ток и воспринимая силовые линии, искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния нескольких метров даже в мутной воде. В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
1. Сильно электрические виды — имеют большие электрические органы, генерирующие разряды от 20 до 600 и даже 1000 В. Основное назначение разрядов — нападение и оборона (электрический угорь, электрический скат, электрический сом).
2. Слабоэлектрические виды — имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов — локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты).
3. Неэлектрические виды — не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10 – 15 м. в морской воде и до 2м. в пресной воде.
Основное назначение генерируемого электричества — локация, ориентация, сигнализация (многие морские и пресноводные рыбы: например, ставрида, атерина, окунь и др.).
Электрические явления имеют собственное значение или сопровождают многие жизненные процессы, если происходит перемещение ионов, а также имеется несимметричное распределение ионов по разные стороны биологических мембран. Это обусловливает наличие электрических потенциалов и возникновение электрических токов. Электрический ток имеет определенное значение в жизни рыб. Он служит для передачи сигналов в некоторых синапсах, приводит к действию отдельные клеточные механизмы. Электрические токи, генерируемые рыбами, позволяют им общаться друг с другом, а также используются для поиска, локации, защиты и нападения.
Природа биотоков и биопотенциалов. Клеточная мембрана отделяет внутреннюю среду клетки, богатую ионами калия, от межклеточной жидкости, богатой ионами натрия. Она окружена снаружи электростатически фиксированными ионами натрия, изнутри — слоем ионов хлора, которые притянуты электростатическими силами к ионам натрия. Ионы калия оттеснены в глубь клетки. Наличие ионной оболочки из положительно заряженных ионов натрия снаружи клетки и ионной подкладки из отрицательно заряженных ионов хлора изнутри мембраны, создает электрическую поляризацию мембраны с разностью потенциала 60—70 мВ в нервных клетках, около 80 мВ в мышечных клетках и 20 мВ в эпителиальных клетках.
Эта поляризация и электрический потенциал поддерживаются низкой проницаемостью мембран для ионов и постоянной работой ионных насосов. Клеточная мембрана обладает высоким электрическим сопротивлением — порядка 1000 Ом/см2 и электрической емкостью около 1 мкФ/см2. Движение ионов по градиенту концентрации и работа ионных насосов порождают ток покоя (рис. ). Токи действия возникают при проведении нервного импульса по нерву при сокращении миофибриллы, при работе клетки почечного канальца, энтероцита и т. д. Специфические электрические явления возникают при освещении светочувствительных клеток, раздражении механорецепторов.
При возбуждающем воздействии на клетку потенциал ее мембраны изменяется. Например, при раздражении нервной или мышечной клетки ее мембрана деполяризуется, т. е. мембранный потенциал уменьшается, а затем возвращается к норме через фазу гиперполяризации. Весь процесс занимает тысячные доли секунды. Сложение токов действия отдельных клеток организма рыб, вызывает возникновение так называемых биотоков – токов возникающих в живом организме, которые обнаруживаются даже на некотором расстоянии от места их возникновения. Запись биотоков работающей мускулатуры называют электромиограммой. По электромиограммесудят о том, как, когда и какие группы мышц работают при различных двигательных актах. Биотоки регистрируются при плавании рыб дыхательных движениях. Электрокардиограмма рыб имеет характерные «зубцы» PQRST. Перед зубцом Р, являющимся результатом сокращения предсердия, у рыб имеется волна возбуждения венозного синуса. Зубец R сопровождает сокращение желудочка, а зубец Т возникает при электрической переполяризации поверхности желудочка.
Для обнаружения тока действия (акционнъй ток), один электрод надо приложить к невозбужденному участку ткани, а другой — к возбужденному и соединить их с регистрирующим прибором.
Записи биотоков сетчатки глаза (ретины) называются ретинограммами. Характерной особенностью их является то, что волна тока действия в них в отличие от мышц и нервов является волной гиперполяризации, а не волной деполяризации. При освещении колбочек и палочек разность потенциалов между внутренней и внешней поверхностями мембраны растет. По характеру сигналов от сетчатки можно определить спектральную чувствительность.
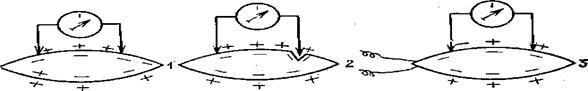
Рис. 74. Мышечные токи:
§
Ток покоя; 2.— ток повреждения; 3 — ток действия
Запись биотоков головного мозга называется энцефалограммой. Как и у других позвоночных животных, энцефалограммы рыб имеют характер коротких скачков и медленных волн. Их изучение может служить для определения действия медикаментов и токсикантов, а также для решения многих научных задач, связанных с изучением нервной деятельности рыб.
При повреждении ткани в участке повреждения анионы выходят на поверхность, и этот участок становится заряженным отрицательно по отношению к другим участкам. Таким образом, если поставить один электрод на поврежденный (отрицательный.) и второй на неповрежденный (положительный) участки, то регистрирующий прибор покажет наличие тока — тока повреждения.
Биотоки возникают при работе органов и тканей у разных животных, но только у рыб имеются специализированные органы генерирующих электрический ток.
Строение электрического органа. Электрические органы представляют собой столбики, состоящие из клеток плоской формы, — электрические пластинки, расположенные по бокам тела. Толщина этих клеток не превышает 10 мкм. Электрические клетки у большинства рыб имеют мышечное происхождение, у электрического сома они происходят из клеток кожи, а у рыб из семейства стернархид — из нервных клеток. Каждую клетку покрывает оболочка — электролемма.
Форма клеток разнообразна: округлая и цилиндрическая (у гимнотид), розеткообразная (у электрического сома) чашеобразная и дискообразная (у скатов), а сторона пластинки к которой подходят нервные окончания, называется лицевой и под обыкновенным микроскопом выглядит гладкой, а под электронным видны канальцы, по которым проходят нервные волокна.

Рис. 75. Расположение электрических органов у рыб: а – скат; б – японский скат; в – электрический сом; г — электрический угорь; д- нильский слоник; е – нильская щука.
Благодаря наличию полупроницаемой клеточной мембраны происходит перераспределение ионов натрия, калия, кальция, хлора. С противоположной стороны клетки подходят кровеносные сосуды, здесь же располагаются и ядра. Эта сторона поверхности у большинства рыб губчатая, морщинистая, и ее называют нутритивным (питательным) слоем.
У электрических рыб различают два вида мембран: возбудимые при помощи химических медиаторов и возбудимые не только химическим, но и электрическим путем. Мембраны первого вида имеются у таких морских рыб, как электрические скаты, звездочеты и др. мембраны второго вида встречаются у пресноводных рыб.
Неровная поверхность пластинки при возбуждении является электроотрицательно по отношению к ее обратной стороне. Исключение составляет электрический сом. Если поставить электроды на разные точки одной стороны пластинки, то регистрирующий прибор не покажет наличия тока, т. е. лицевая сторона работает как единое целое. Если один электрод поставить на лицевую сторону пластинки, а другой погрузить в ее цитоплазму или поставить на противоположную сторону, то возникает ток напряжением 90—150 мВ.
Контрольные вопросы.
1.Мофологические особенности плавников костистых и хрящевых рыб?
2.Функциональное значение плавников для жизнедеятельности рыб?
3.Каков механизм движения рыб при плавании?
4.Охарактеризуйте механизм полета рыб.
ГЛАВА 11
Центральная нервная система выполняет в организме функции первостепенной важности. Во-первых, она обеспечивает точную регуляцию всех процессов внутри организма, их координацию и интеграцию, благодаря чему все органы и системы действуют согласованно, а организм представляет собой единое функциональное целое. Во-вторых, ей принадлежит функция связи организма с постоянно меняющейся внешней средой.
11.1 Общая физиология центральной нервной системы. Центральная нервная система позвоночных построена из громного количества нейронов, вступающих в тесную связь друг с другом. Однако каждый нейрон относительно самостоятелен. Это обусловливается тем, что протоплазма одной клетки и ее отростков не проникает внутрь другой клетки. Аксон одной нервной клетки, как правило, только соприкасается с дендритами или телом другой. Место контакта двух нейронов называют синапсом. В центральной нервной системе имеется бесчисленное множество синапсов.
В функциональном отношении нейроны делят на три основных класса: 1) чувствительные, или афферентные, 2) двигательные, или эфферентные, 3) вставочные, или промежуточные.
Афферентные нейроны проводят возбуждение от рецепторов в центральную нервную систему, их называют также рецепторными. Тела этих нейронов расположены вне центральной нервной системы и находятся в спинномозговых или черепномозговых ганглиях. Данные нейроны отличаются от других наличием двух длинных отростков — собственно аксона, передающего возбуждение от тела клетки в центры спинного мозга или мозгового ствола, и аксоноподобного дендрита, уходящего на периферию в виде афферентного волокна и ветвящегося там на чувствительные нервные окончания — рецепторы. К рецепторным нейронам относятся также некоторые нейроны в центральной нервной системе, которые получают возбуждение не непосредственно от рецепторов, а через другие истинно рецепторные нейроны. К ним относятся, например, нейроны зрительных бугров.
Рецепторные нейроны называют также чувствительными, или сенсорными, так как они доставляют в центральную нервную систему импульсы, вызывающие различные ощущения.
Эфферентные, или эффекторные, нейроны передают возбуждение из центральной нервной системы к рабочим органам — эффекторам. От тел этих нейронов возбуждение идет на периферию по длинным аксонам. Те эфферентные нейроны, которые посылают импульсы к скелетным мышцам, называются двигательными нейронами, или мотонейронами. Их тела лежат в вентральных рогах спинного мозга, в продолговатом и среднем мозге.
Многие эфферентные нейроны передают импульсы не прямо на периферию, а через другие, ниже расположенные нейроны. Например, эфферентные нейроны среднего мозга посылают импульсы к мотонейронам спинного мозга. Эфферентные нейроны вегетативной нервной системы находятся вне центральной нервной системы — в периферических ганглиях.
Вставочные (промежуточные, контактные нейроны, интернейроны) образуют самую многочисленную группу нейронов в центральной нервной системе. Они осуществляют связь между рецепторными и эффекторными нейронами. По характеру вызываемого ими эффекта контактные нейроны делят на возбуждающие и тормозящие.
11.2Строение и классификация межнейронных синапсов.Межнейронным, или центральным, синапсом называют место контакта окончания аксона с телом или отростками другой нервной клетки. Если аксон оканчивается на теле (соме) другой нервной клетки, синапс будет аксосоматическим, на дендритах — аксодендритическим, на аксонах-аксоаксональным. Синапсы состоят: 1) из пресинаптической мембраны, покрывающей окончание аксона (синаптическую бляшку), 2) синаптической щели шириной примерно 200—ЗООА и 3) постсинаптической мембраны соседней нервной клетки. Внутри синаптической бляшки находится много пузырьков, в которых, как полагают, содержится вещество, обеспечивающее передачу импульса через синапс. Это вещество получило название медиатора. Кроме пузырьков медиатора, имеются митохондрии — специфические структуры, поставляющие клетке энергию. По функциональному признаку различают возбуждающие и тормозящие, или тормозные, синапсы. Возбуждающие и тормозящие синапсы имеют некоторые структурные отличия. Так, возбуждающие синапсы большей частью являются аксодендритическими, характеризуются относительно широкой синаптической щелью (примерно ЗООА), толстой, плотной постсинаптической мембраной. В синаптической щели могут быть специальные включения в виде пластинок внеклеточного вещества. Пузырьки медиатора относительно крупные, округлой формы. Тормозные синапсы имеют более узкую синаптическую щель (в 1,5—2 раза уже, чем возбуждающие); постсинаптическая мембрана более тонкая; в синаптической щели нет включений внеклеточного вещества; синаптические пузырьки овальной формы, по размеру меньше, ряд пузырьков уплощен.
Природа медиаторов возбуждающего и тормозного синапсов также различна. Химическим передатчиком возбуждения в возбуждающих синапсах является ацетилхолин; полагают, что такими свойствами обладает и глутаминовая кислота. В возбуждающих центральных синапсах медиаторами служат адреналин, норадреналин, 5-окситриптамин (близкие по строению вещества), а также гистамин.
К медиаторам, выделяющимся в тормозящих синапсах, относят аминокислоту глицин. Глицина в участках спинного мозга, где распола
-гаются тормозные нейроны Реншоу, значительно больше, чем в других соседних местах. Кроме того, когда методом электрофореза к телу нерв
ной клетки, подводили глицин, возникала — гиперполяризация клеточной мембраны, что является доказательством постсинаптического торможения. Вторым тормозящим медиатором считают гамма-аминомаслянуго кислоту (ГАМК). Этот медиатор был открыт у ракообразных. Затем он был найден в головном мозге в частности в окончаниях тормозящих нейронов. Механизм передачи возбуждения в синапсах центральной нервной системы.Создание тонких методов регистрации импульсов отдельных нейронов с помощью высокочувствительных электрических приборов произвело переворот в исследованиях передачи импульсов через синапс. Этот метод заключается в том, что в нервную клетку осторожно вводят очень тонкую стеклянную пипетку, заполненную солевым раствором, проводящим электрический ток (например, концентрированным раствором хлористого калия). Если ввести осторожно пипетку и закрепить ее неподвижно, клеточная оболочка быстро стягивается вокруг кончика пипетки, и такие нервные клетки способны нормально функционировать в течение нескольких часов.
Механизм передачи возбуждения заключается в следующем. Под действием приходящих к окончанию аксона нервных импульсов происходит выделение из синаптических пузырьков молекул медиатора ацетилхолина. Они диффундируют через пресинаптическую мембрану в синаптическую щель, а затем к постсинаптической мембране и прикрепляются к определенным ее участкам, называемым рецепторными. Здесь медиатор взаимодействует с ее структурными компонентами—белково-липоидными комплексами. В результате происходит кратковременное увеличение диаметра пор в мембране (в 3—4 раза), что приводит к сильному повышению проницаемости постсинаптическоймембраны для ионов натрия, которые устремляются из синаптической щели через постсинаптическую мембрану. Происходит деполяризация постсинаптической мембраны и возникает возбуждающий постсинаптический потенциал (ВПСП), и когда он достигает критического уровня, появляется потенциал действия. Выделившийся ацетилхолин очень быстро разрушается ферментом холинэстеразой на холин и уксусную кислоту. Поэтому каждая выделившаяся порция медиатора действует кратковременно, успевая вызвать только один импульс. Сразу после, возникновения импульса происходит уменьшение диаметра пор в постсинаптической мембране, и ионы натрия, которые в гидратированном состоянии крупнее ионов калия, перестают переходить внутрь клетки, а ионы калия свободно выходят наружу в синаптическую щель. Благодаря этому всего лишь за милли секунду или даже быстрее восстанавливается исходная полярность постсинаптической мембраны и включается натриево-калиевый насос (рис. 76).
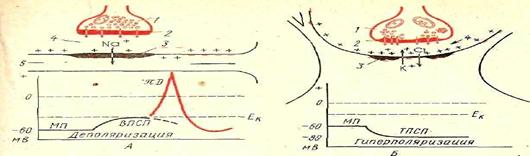
Рис76. Схема функции синапса:
А — возбуждающего; Б — тормозящего; 1 — пузырьки медиатора; 2 — лресинаптическая мембрана; 3 — постсинантическая мембрана; 4 — синаптическая щель; Ек — критический уровень деполяризации; МП — местный потенциал; ВПСП — возбуждающий и ТПСП — тормозной посчеинаптические потенциалы; ПД — шненциал действия.
11.3Учение о рефлексе.Основной формой деятельности центральной нервной системы является рефлекс.
Рефлексом называют реакцию организма на раздражение рецепторов, осуществляемую при участии центральной нервной системы.
Учение о рефлексе имеет свою историю, охватывающую столетия. Основоположником представлени о простейших рефлекторных актах является французский философ и естествоиспытатель Рене Декарт(1595—1650). Он описал такие автоматические действия, как отдергивание руки при прикосновении к огню, подмигивание глаз при раздражении роговицы и т. д. Большую роль в дальнейшем развитии учения о рефлексе сыграл чешский анатом и физиолог Георг Прохаска, который и ввел в науку этот термин. Слово «рефлекс» в переводе с латинского значит «отражение», т. е. в него вкладывался такой смысл, что всякое действие организма является отражением какого-либо воздействия на него и для каждого действия имеется своя причина. Представление о рефлексе как о явлении, впервые сделал русский ученый-материалист И. М. Сеченов в своей знаменитой книге «Рефлексы головного мозга» (1863), обосновавший положение о том, что «все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». И. П. Павлов, развивая и продолжая идеи И. М. Сеченова, создал учение об условных рефлексах и применил метод условных рефлексов для исследования функций головного мозга.
Понятие о рефлексе и рефлекторной дуге. Морфологической единицей нервной системы считается нейрон, а функциональной единицей нервной деятельности является рефлекс. Рефлекторные акты представляют собой основную специфическую функцию нервной системы.
Прежде чем рассматривать деятельность отдельных частей центральной нервной системы, необходимо рассмотреть основные закономерности рефлекторных реакций.
Под рефлексом понимают непроизвольную ответную реакцию организма на раздражение, осуществляемую центральной нервной системой и выражающуюся мышечной или секреторной деятельностью.
Любая рефлекторная реакция осуществляется через так называемую рефлекторную дугу, которая состоит из двух и более нейронов (рис.77а). Рефлекторная дуга это путь, по которому проходят нервные импульсы от рецептора до эффектора. Сущность рефлекторного акта состоит в последовательном возбуждении различных участков рефлекторной дуги. Раздражение внешней среды воспринимает рецептор, т.е.окончания центростремительного нерва в коже (экстерорецептор), в слизистых оболочках (интерорецептор) и в других частях организма.
Возникший в результате раздражения рецептора процесс возбуждения передается через центростремительный (афферентный) нерв в чувствительную клетку, а от неё, через дендрит,- двигательной клетке серого вещества спинного мозга. Далее импульс возбуждения идёт по центробежному (эфферентному) нерву к рабочему органу — эффектору,т. е. к мышце или железе.
Рис.76 а
А — спинной мозг; В — двигательный (нижний) корешок, С—чувствительный (верхний) корешок; D — спинномозговой узел; Е—кожа, F—мышца скелета, о — клетки чувствительных нейронов в спинномозговых узлах, Ь — клетки двигательных нейронов в передних рогах, с — периферические отростки чувствительных нейронов, которые, оканчиваясь в коже, образуют рецепторы d—нейрит двигательных нейронов, оканчивающихся в скелетных мышцах.
В осуществлении рефлекторной функции может участвовать вся центральная нервная система.
Необходимым условием рефлекторного процесса является целость всех составляющих частей дуги. Нарушение одной из этих частей, например, рецептора, делает невозможным осуществление рефлекса через данную дугу.
Для животных и рыб рефлекторные акты имеют большое приспособительное значение. Выключение некоторых рефлекторных функций центральной нервной системы может значительно нарушить отправление важнейших для жизни частей организма.
Рефлекторные акты осуществляются через нервные клетки различных этажей центральной нервной системы. При этом возбуждение по многочисленным проводящим путям способно широко распространяться в центральной нервной системе. Рефлекс — это сложная реакция организма, осуществляемая нервными центрами. Поэтому необходимо обратить внимание на некоторые основные свойства центров, от которых в значительной степени зависит конечный эффект рефлекторного акта. Основные свойства нервных центровОдностороннее проведение. У позвоночных животных импульс в центральной нервной системе проводится только в одном направлении: от рецепторного нейрона через промежуточный к эффекторному. Такая направленность нервного процесса, характерная для рефлекторной дуги, обусловлена односторонним проведением возбуждения в синапсах. Такое явление можно объяснить тем, что синапс представляет собой односторонне проницаемую перепонку.
Замедленное проведение. Проведение возбуждения в центральной нервной системе совершается значительно медленнее, чем по нервам. Это объясняется свойствами синапсов задерживать проведение возбуждений с одного нейрона на другой. Проведение через синапс у лягушки осуществляется в течение 0,001 —0,003 секунды. Очевидно, чем большее число нейронов входит в данную рефлекторную реакцию, тем больше затрачивается времени на её осуществление.
Временем рефлекса называется период, протекающий от момента раздражения до начала реакции. Например, время рефлекса моргания у человека составляет 0,05—0,2 секунды.
Облегчение. Раздражение, приходящее в нервные центры, не исчезает сразу, а оставляет след в виде повышенной возбудимости блуждающего нерва.
Торможение.В центральной нервной системе, наряду с возбуждением, встречается другой процесс — торможение. Благодаря тесной функциональной взаимосвязи возбудительного и тормозного процесса, а также взаимодействия между отдельными нейронами, называемого координацией, обеспечивается наиболее точное и тонкое мышечное движение и приспособление тех или иных двигательных актов к тем или иным воздействиям внешней среды.
Впервые процесс центрального торможения был открыт Сеченовым. Для возникновения торможения необходимо одновременное поступление в центральную нервную систему двух или больше импульсов возбуждения по различным рефлекторным путям. Более сильное раздражение вызывает угнетение аффекта от более слабого раздражения. Объясняется это тем, что в центральной нервной системе происходит конфликт между возбуждениями, и одно из них подавляет другое. Так, например, раздражение серого вещества головного мозга может повести к задерживанию спинных рефлексов и т. д. Природа процесса торможения ещё не ясна.
Рефлекторное сокращение мышц сгибателей ведёт к расслаблению мышц разгибателей той же конечности. Мышцы, производящие сгибание конечности в одном или нескольких суставах, называются синергистами, а мышцы, производящие разгибание,т.е. противоположное действие, называются антагонистами.
Взаимоотношения между мышцами антагонистами в период любого двигательного акта обусловлены соотношением нервных процессов в центрах. Возбуждение центров антагонистических групп мышц возникает в момент сокращения и обусловлено восприятием раздражения рецепторами, заложенными в мышцах и сухожилиях; эти рецепторы называются проприорецептор а м и.
По имеющимся данным, возбуждение центра одной группы мышц связано с торможением центра другой, антагонистической, мускульной группы (так называемый принцип взаимносопряжённой, реципрокной иннервации). Реципрокные соотношения между центрами осуществляются в широких пределах. Так, например, возбуждение центра сгибания одной конечности вызывает не только торможение центра разгибания этой же конечности, но и торможение центра сгибания, и возбуждение центра разгибания мышц другой конечности. Вследствие этого, при сгибании одной конечности происходит разгибание другой.
Реципрокные взаимоотношения имеются также между центрами передних и задних конечностей. В центрах мускулатуры конечностей, участвующих при двигательных актах, постоянно происходит перераспределение возбуждения и торможения.
г) Изменчивость координации и доминанта
Изучение взаимоотношений между центрами, иначе — координационных отношений, показывает, что имеются относительно постоянные, сложившиеся в процессе фило- и онтогенетического развития явления координации, как, например, реципрокная иннервация антагонистических мышц. Но в нервной системе мы наблюдаем и изменчивость явлений координации. Координационные отношения складываются в результате целого ряда внутренних и внешних условий к моменту возникновения данного раздражения. Ответная рефлекторная реакция может поэтому значительно видоизменяться.
Рис.77. Схематическое изображение реципрокной
иннервации.
Раздражение чувствительного нерва вызывает на соответствующей стороне возбуждение двигательного нейрона, принадлежащего сгибателю- F; одновременно вызывается торможение двигательного нейрона — е, принадлежащего разгибателю, раздражение на другой стороне производит противоположный эффект.
В связи с динамичностью координационных процессов особый интерес представляет установленный Ухтомским принцип доминанты, позволяющий понять механизм преобладания одних очагов возбуждения над другими.
Доминанта — это господствующий очаг возбуждения в центральной нервной системе. Появляется такой очаг при условии повышения возбудимости какого-либо центра рядом волн возбуждения, оптимальных по ритму и силе или по гуморально — химическим влияниям. Доминантный очаг возбуждения привлекает к себе возбуждения, приходящие к другим нервным центрам, усиливается за счёт этих возбуждений и затормаживает все остальные центры.
11.4 Морфология и функциональная характеристика центральной нервной системы.Центральная нервная система рыб, как и у ланцетника, имеет вид трубки. Ее задний отдел — спинной мозг — расположен в канале позвоночника, образованном верхними дугами позвонков. От спинного мозга между каждой парой позвонков вправо и влево отходят нервы, управляющие работой мышц тела и плавников. По нервам от чувствительных клеток на теле рыбы в спинной мозг поступают сигналы о раздражении. Передняя часть нервной трубки рыбы и других позвоночных животных видоизменена в головной мозг, защищенный костями черепной коробки. В головном мозге позвоночных различают отделы: передний мозг, промежуточный мозг, средний мозг, мозжечок и продолговатый мозг. Все отделы головного мозга имеют большое значение в жизнедеятельности рыбы. Например, мозжечок управляет координацией движений и равновесием животного. Продолговатый мозг постепенно переходит в спинной мозг. Он играет большую роль в управлении дыханием, кровообращением, пищеварением и другими важнейшими функциями организма
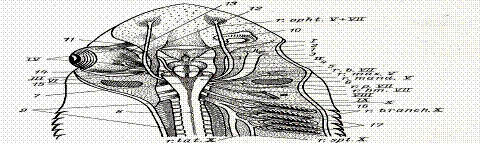
Рис.79. Головной мозг и головные нервы колючей акулы (со спинной стороны, правый глаз удален):
1- предний мозг, 2 – обонятельная доля переднего мозга, 3 – промежуточный мозг, 4 – средний мозг, 5 — мозжечок, 6 – продолговатый мозг, 7 – ромбовидная ямка, 8 – спинной мозг, 9 – спинномозговые нервы, 10 – обонятельный мешок, 11 – верхняя косая мышца глаза, 12 – верхняя прямая мышца глаза, 13 -внутренняя прямая мышца глаза , 14 — наружная прямая мышца глаза , 15 — брызгальце, 16 – первая жаберная щель, 17- вторая – пятая жаберные щели; I-обонятельный тракт; I I – зрительный нерв; III – глазадвигательный нерв; IV- блоковый нерв;V-тройничный нерв;верхнечелюстная ветвь; нижнечелюстная ветвь; VI — ,VII-, VIII- ,IX-, X-, X -,X —
Нервная система делится на центральную, к которой относятся головной и спинной мозг, и периферическую – нервы, отходящие от головного и спинного мозга. От мозга отходят нервные волокна, окончания которых выходят на поверхность кожи и образуют у большинства рыб ярко выраженную боковую линию, проходящую от головы до начала лучей хвостового плавника. Боковая линия служит для ориентации рыбы:определения силы и направления течения, наличия подводных предметов 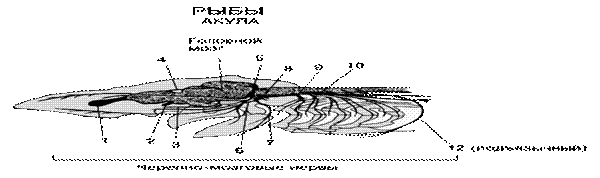
Рис.79. Расположение черепномозговых нервов акулы
Наблюдая поведение окуня в аквариуме, можно заметить, что ответы на раздражение у окуня могут проявляться двояко. Если к окуню прикоснуться, он моментально метнется в сторону. Столь же быстр его ответ на вид пищи. Жадный хищник, он стремительно кидается на свою добычу (мелких рыб и различных беспозвоночных — ракообразных, червей). При виде добычи возбуждение идет по зрительному нерву в центральную нервную систему окуня и сейчас же по двигательным нервам возвращается от нее к мускулам. Окунь плывет к жертве и захватывает ее.
Механизм подобных ответов организма на раздражение врожденный. А такие рефлексы называют безусловными. У всех животных одного вида безусловные рефлексы одинаковы. Они врожденны и передаются по наследству.
Если кормление рыб в аквариуме сопровождается какими-либо действиями (условиями), например зажиганием лампочки или постукиванием по стеклу, то через некоторое время такой сигнал начинает привлекать рыб сам по себе, без подкормки. На подобные сигналы у рыб вырабатываются условные рефлексы, то есть рефлексы, возникающие при определенных условиях. В отличие от инстинктивных безусловных рефлексов условные рефлексы не передаются по наследству. Они индивидуальны и вырабатываются в течение жизни животного.
Головной мозг: У акулы головной мозг состоит из пяти отделов. Передний мозг (telencephalon) крупный, нечетко разделен на правую и левую (полушарии). В передней части полушарии хорошо заметны сильно развитые обонятельные части.
Задняя часть переднего мозга незаметно переходит в промежуточный мозг (diencephalon), который на препарате виден, как углубление между вздутиями переднего и среднего мозга. Средний мозг (mesencephalon) имеет вид крупных парных вздутий (зрительные доли), которые прикрывают собой остальную массу этого отдела мозга.
Четвертый отдел мозга – мозжечок (cerebellum, metencephalon) у акуловых рыб сильно развит и спереди налегает на средний, а сзади на продолговатый мозг.
Последний отдел головного мозга – продолговатый мозг (myelencephalon). Сверху в центральной части продолговатого мозга отчетно видна ромбовидная ямка, представляющая собой полость этого отдела мозга (четвертый желудочек). Она затянута сосудистой оболочкой, которая при препарировании обычно разрушается.
Головные нервы. Характер отхождения головных нервов у акуловых рыб типичен для всех позвоночных животных. От головного мозга у акуловых рыб отходят десять пар головных (черепных) нервов, каждая пара симметрична с обеих сторон головного мозга. Головные нервы обычно имеют двойное обозначение: порядковый номер и название.
I. Обонятельный нерв (nervus olphactorius). У акуловых рыб, как и у всех позвоночных, от переднего мозга отходят выросты его лобных долей – обонятельные тракты. Они образуют у обонятельных капсул расширение – обонятельные луковицы. Обонятельный нерв представляет комплекс коротких чувствительных волокон, идущих от эпителия обонятельного мешка к обонятельной луковице.
II. Зрительный нерв (n. opticus) фактически это зрительный тракт, образующийся как вырост стенки мозга он отходит ниже боковой поверхности промежуточного мозга и пройдя отверстие в стенке глазницы, входит в глазное яблоко, распространяясь по внутренней поверхности сетчатки. Нерв чисто чувствительный.
У всех позвоночных животных движения глазного яблока осуществляется при помощи глазных мышц, каждая из которых одним концом прикрепляется к стенке глазницы, а другим к определенному участку поверхности глазного яблока. От передней части стенки глазницы, отходят 2 косые мышцы глаза; верхняя (m. Obliquus superior ) и нижняя (m. Obliquus interior). От задней стенки глазницы пучком отходят 4 прямые мышцы глаза: прямая нижняя (m. Recfus inferior), прямая внутренняя (m. Rectus internus), прямая верхняя (m. Rectus zuperior) и прямая наружная (m. Rectus externus). Эти мышцы иннервируются тремя парами головных нервов – III, IV u VI все эти нервы чисто двигательные.
Глазодвигательный нерв (n. oculomotorius). Отходит от дна среднего мозга, пронизывает стенку черепа и делится у основания прямых мышц глаза на ветви, идущие к четырем мышцам к нижней косой и к нижней внутренней и верхней прямым.
Блоковый нерв (n. trochlcaris). В виде тонкой нити отходит от задне-верхней части среднего мозга, проходит вперед по крыше среднего моста, и проидя через переднюю стенку глазницы, разветвляется в верхней косой мышце глаза. Все остальные головные нервы отходят от продолговатого мозга.
Отводящий нерв (n. abolucens). От дна продолговатого мозга сразу уходит в дно черепной коробки. Он иннервирует наружную прямую мышцу глаза.
Тройничный нерв (n. trigeminus). Это нерв сложный. Он отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу же делится на три ветви глазничную, верхнечелюстную и нижнечелюстную. Глазничная ветвь (ramus ophtholmicus) вместе с глазничной ветвью лицевого нерва проходит через глазницу и ветвится в передней части рыла, иннервируя кожные органы чувств.
§
Верхнечелюстная (r. maxillaris) и нижнеечелюстная (r. manolibularis) ветви, начинаются общим стволом и обособляются позади глаза, они иннервируют мышцы челюстной дуги, зубы, слизистую оболочку ротовой полости. По функции, тройничный нерв смешанный, он имеет чисто чувствующую ветвь-глазничную и смешанные (чувствующие и двигательные) верхнечелюстную и нижнечелюстную.
Лицевой нерв (n. facialis). Отходит сразу же за тройничным. Этот нерв также распадается на несколько ветвей наиболее крупные из них — глазничная, щечная, небная и подъязычная ветви.
Глазничная ветвь (r. ophtalmicus) идет вместе с одноименной ветвью тройничного нерва. Позади глазничной ветви отходит крупный ствол щечная ветвь (r. buccalis). Она в виде широкого тяжа отходит по дну глазницы, иннервируя кожные органы чувств брюшной поверхности головы. Далее общим стволом отходят две ветви: небная и подъязычная.
Небная ветвь (r. palatinus) сразу же после обособления уходит в дно черепной коробки и затем ветвится в слизистой оболочке ротовой полости.
Подъязычная ветвь (r. hyamanolibularis) проходит позади брызгальца и иннервирует мускулатуру подъязычной дуги и кожные органы чувств боков головы. Лицевой нерв, как и тройничный смешанный, его глазничная, щечная и небная ветви чувствующие, а подъязычная смешанная (чувствцющая и двигательная).
Слуховой (n. acusticus). Идет от боковой поверхности продолговатого мозга и почти сразу же входит в стенку черепной коробки, иннервируя внутреннее ухо. Это чисто чувствующий нерв.
Языкоглоточный нерв (n. glozsopharynqeus). Подходит к первой жаберной щели и здесь делится на две ветви иннервирующие соответственно ее переднюю и заднюю поверхности. Задняя ветвь иннервирует всю мускулатуру первой жаберной дуги. Этот нерв по своей функции смешанный, он имеет и чувствующие и двигательные волокна.
Блуждающий нерв (n. vagus). Отходит от задне — боковой поверхности продолговатого мозга несколькими корешками почти сразу же сливающимися в толстый нервный тяж. Иннервирует обширную область тела, его основные ветви — четыре жаберные, внутренностная и боковая.
Четыре жаберные ветви (r. branchialis) иннервируют жаберные щели от второй до пятой. Каждая ветвь пройдя к жаберной щели, делится на две передне — и заднежаберную. Переднежаберная ветвь чувствующая и заднежаберная смешанная вместе иннервируют всю мускулатуру, соответствующей жаберной дуги (от второй до пятой).
Внутренностная ветвь (r. splanchnicus) идет как продолжение основного ствола блуждающего нерва. Она входит в брюшную полость, иннервируя внутренние органы. Поверхностно расположена боковая ветвь (r. lateralis) иннервирующая органы боковой линии туловища и хвоста. Блуждающий нерв смешанный (включает и чувствующие, и двигательные волокна). У высших позвоночных животных обособляются еще две пары головных нервов.
Подъязычный нерв (n. hypoqlozzus) впервые четко обособляется у пресмыкающихся. Зачатки этих нервов есть у акуловых рыб в виде нервных корешков, отходящих позади блуждающего нерва в области затылочной стенки мозгового черепа.
Спинномозговые нервы (n. spinalis) посегментально отходят от спинного мозга (medulla spinalis). Иннервирует, соответствующие сегменты тела и по своей функции представляют собой смешанные нервы, состоящие из чувствующих и двигательных волокон.
Органы боковой линии. Органы боковой линии представляют собой сформировавшуюся в процессе эволюции высокоспециализированную сенсорную систему, существующую у первичноводных позвоночных животных, круглоротых, рыб. Органы боковой линии расположены в коже и подкожных структурах, однако по своему строению они сходны с рецепторами внутреннего уха. В эмбриональный период оба этих типа рецепторов развиваются из одного и того же эктодермального утолщения височной, области, т.е. имеют общее происхождение, орган боковой линии выполняет четко выраженную механорецепторную функцию.
Адекватным стимулом для этой системы являются смещения водной среды, возникающие при различных колебаниях водной среды, перепадах давления в потоках воды, в том числе и при действии низко частичных звуковых волн.
В составе боковой линии некоторых рыб наряду с механорецепторными структурами существуют и нижесенсорные образования, предназначенные для восприятия слабых электрических полей водной среды. Для этого чтобы различать эти два типа сенсорных приборов, механорецепторные органы стали называть обычными, а электрорецепторные образования — специализированными.
Источниками сигналов для электроценторов могут служить самые разнообразные электрические явления, возникающие в водной среде в результате различных физических и физико — химических процессов, а также биологическими процессами, связанными с жизнедеятельностью животных.
Всех рыб, обладающих развитым электрорецепторным аппаратом, в соответствии с их способностью к генерации электрических полей принято подразделять на 3 группы;
1) группу составляют неэлектрические рыбы, не имеющие электрических органов. К ним относятся большинство пластиножаберных, за исключением семейства обыкновенных скатов, а также ряд представителей отряда сомовых, например карликовый сомик, туркестанский сомик.
2) группу составляют слабоэлектрические рыбы, их электрические органы генерируют разряды небольшой величины, не превышающие по амплитуде 10-15В. В эту группу входят, представители подотряда электрических угрей, обитающих в реках Южной Америки, представители отряда мормирообразных, включающих два подотряда – гимнархид и мормирид, живущих в африканских реках, и представители семейства обыкновенных скатов.
3) К группе относятся сильноэлектрические рыбы, способные генерировать как низковольтные, так и высокоамплитудные разряды, достигающие нескольких сотен вольт.
Механорецепторные структуры боковой линии. Основу органов боковой линии представляют специализированные волосковые клетки — нейромасты (рис. ), располагающиеся на основной мембране. Каждый нейромаст имеет различные виды волосков: один крупный — киноцилий и пучок из 40— 50 мелких — стереоцилий. Волоски погружены в желеобразную среду — купулу. Несколько нейромастов объединяются и образуют чувствительный бугорок. Кроме того, имеются опорные эпителиальные клетки.
Наиболее просто органы боковой линии устроены у круглоротых и некоторых костистых рыб (колюшка), у которых они представлены продольными рядами чувствительных клеток, расположенных на поверхности тела,— поверхностные органы. У акул и химер нейромасты расположены на дне желобка, проходящего по бокам тела, у ганоидов и большинства костистых рыб эти клетки располагаются на дне канала, сообщающегося с поверхностью тела небольшими тонкими ходами. Это канальные органы.
У многих рыб органы боковой линии располагаются не только по бокам тела, но и в области головы, на жаберных крышках и нижней челюсти. Степень развития боковой, линии у разных видов рыб различна. У верховки и корюшки она распространяется на небольшое расстояние позади головы, у других рыб она хорошо развита на всем протяжении туловища.
У костистых рыб и амфиогии одиночный эпидермальный орган, или невромаст состоит из 20-60 рецепторных клеток, снабженных усилиями или волосками окружающих их опорных клеток и желатинообразной купулы, покрывающей сверху волосковые клетки.Совокупность рецепторных и опорных клеток образует сенсорный эпителий. Волосковая клетка имеет длину в среднем 30-40 и ширину 7-10 мкм. Ее цилиарный аппарат состоит из нескольких десятков стереоцилии и одной киноцилии, длинной около 40 мкм. Стереоцими и киноцилия своими дистальными концами погружены в капсулу, которая у свободных невромастов может выступать над поверхностью кожи до 100 мкм.
Вследствие того, что киноцилия располагается всегда асимметрично относительно пучка стереоцилии, рецепторная клетка оказывается морфологически поляризованной. У рыб одиночная волосковая клетка иннервируется обычно несколькими первичными волосками, образующими на базальной части клетки окончания двух типов: гранулированные и негранулированные органы боковой линии иннервируются передним и задним нервами боковой линии. Передний нерв снабжает рецепторные структуры на голове, а задний на туловище животного. Нервы боковой линии представляют собой периферические отростки ганглиозных клеток, лежащие внутрикраниально. Передний нерв боковой линии образован ветвью лицевого или тройничного нервов, а задний ветвями блуждающего и языкоглоточного нервов.
Механорецепторные структуры боковой линии представлены двумя разновидностями чувствительных образований, свободными невромастами и канальными органами. Основное отличие между ними заключается в особенностях структуры вспомогательного аппарата, благодаря которому свободные невроматы непосредственно контактируют с наружной средой, а канальные органы такого контакта не имеют и внешний стимул вызывает, прежде всего, перемещение канальной жидкости, которое и воспринимается рецепторами. У костистых рыб сенсорные каналы состоит из 5 основных ветвей: главного латерального канала, надвисочного, височного, над и подглазничного каналов.
Функциональные свойства электроцепторной структуры.
Локальные электрические явления в рецепторах. В рецепторах боковой линии, как и в других вторичночувствующих рецепторах, функцию генераторного потенциала выполняет возбуждающий постсинаптический потенциал (ВПСП). Частое ограничение системы осуществляется не на уровне рецепторов, а за счет механических свойств окружающих их элементов, взаимодействующих с водной средой.
Синаптическая передача. Между волосковой клеткой и окончаниями сенсорного нейрона имеются синоптические контакты, характерные для синапсов с химическим механизмом передачи.
Эфферентная регуляция. Эфферентные влияния на деятельность механорецепторов боковой линии носят тормозной характер и воздействуют на спонтанную и вызванную активность рецепторов.
Эфферентные влияния, снижая общий уровень фоновой активности рецепторов, уменьшают не только фоновый шум, существующий в данной сенсорной системе, но и величину рецептивного поля, что имеет важное значение в локализации источника стимула в пространстве.Эфферентные импульсы в органах боковой линии рыб могут быть вызваны афферентными разрядами, причем таким действием обладают также и контралатеральные афферентные входы. В эфферентной системе в качестве медиатора выделяется ацетилхолин.
Импульсивная активность. В состоянии покоя в нервных волокнах, иннервирующих невромасты бок линии, регистрируется довольно высокий уровень фоновой активности, достигающий 100 имп./сек. наличие фоновой активности в нервных волокнах отражает фоновую активность рецепторов и связано со спонтанным выделением медиатора из волосковых клеток.
Специализированные рецепторы боковой линии отличаются большим разнообразием. Наиболее удачная – это классификация Сабо, согласно которой все электрорецепторы следует подразделять на ампулированные и бугорковые. В ампулированных органах рецепторные клетки сообщают с наружной средой при помощи специального канала, тогда как бугорковые органы такого канала не имеют. Ампулированные рецепторы имеются у всех морских и пресноводных рыб, обладающих электроцепторным аппаратом. Бугорковые рецепторы встречаются только у слабоэлектрических рыб.
Ампулированные рецепторы. К ним относятся не имеющие специального наименования ампулированные органы слабоэлектрических рыб, ампулы Лоренцини пластиножаберных рыб и малые ямковые органы сомовых рыб. Отдельный орган состоит из канала, идущего от поверхности кожи и заканчивающегося слепым расширением — ампулой, в основании которой располагается сенсорный эпителии. У слабоэлектрических рыб длина канала варьирует от 100 до 200 мкм.
Ампулярные органы распределены по всей поверхности тела, но их концентрация в области головы особенно велика. Иначе организованы ампулы Лоренцини, встречающиеся у пластиножаберных рыб и единственной костистой рыбы Plotosus. Длина каналов у этих рецепторов в зависимости от среды обитания варьирует от 500 мкм. У пресноводного ската Potamotrygon до нескольких десятков сантиметров у крупных морских скатов.
Каналы расположены под кожей и имеют различную длину, так как берут начало в различных участках тела и сходятся к капсулам, где заканчиваются ампулярным расширением, на дне которого расположены чувствительные клетки. На одной рецепторной клетке может быть несколько синапсов, а одно и то же нервное окончание может контактировать сразу с двумя соседними рецепторными клетками.
Ампулярные каналы заполнены желеобразным веществом, состоящим главным образом из мукополисахаридов. Рецепторы апмулированных образований функцианируют в условиях окружения некоторой специфической среды, отличающейся повышенным содержанием ионов калия по сравнению с наружной средой и жидкостями тела. Особенности ампулярного канала придают ему свойства идеального морского кабеля, обеспечивающего прохождение сигнала вдоль него почти без ослабления.
Бугорковые рецепторы располагаются в инвагинациях эпидермальной мембраны. Отдельные органы содержат рецепторные клетки, находящиеся в эпидермальной капсуле, не имеющей прямого контакта с наружной средой количество рецепторных клеток в капсуле варьирует от 1 до 35 у мормирид и от 10 до 100 у гипнотид.
Функциальные свойства. Ампулированные рецепторы разных типов имеют принципиально сходное строение, характеризующееся наличием ампулоподобного органа в качестве основной структурной единицы.
Ампулированные рецепторы слабоэлектрических костистых рыб менее чувствительны к электрическим полям. Исследования электрической чувствительности бугорковых рецепторов слабоэлектрических рыб показывают, что этот класс рецепторов оказывается менее чувствительным к электрической стимуляции, чем ампулированные рецепторы.
Ампулированные рецепторы наряду с высокой электрической чувствительностью обладают сильно развитой чувствительностью к изменению температуры окружающей среды. Снижение температуры воды вызывает временное повышение частоты импульсации, а при повышении температуры наблюдаются противоположные изменения активности.
Термочувствительной является область сенсорного эпителия. Механическая чувствительность ампулированных рецепторов выражена слабо. В отношении химической чувствительности рецепторов ампул Лоренцини (восприятие изменении солености, содержание различных ионов, углекислого газа) обусловлена возникновением соответствующих полей электрохимического происхождения.
Центральные структуры системы боковой линии:Продолговатый мозг. Нервы боковой линии оканчиваются в ядрах акустико — латеральной области (area acousticolateralis). Акустика — латеральная область, продолговатого мозга является неоднородным образованием и состоит из нескольких ядер. У гонадно — и поперечноротых рыб ее принято подразделять на дорсальное, медиальное и вентральное ядра. Вентральное ядро связано со слуховой системой. Медиальное ядро, образованная клетками дорсолатеральной части продолговатого мозга, получает волокна, как от переднего, так и от заднего нервов боковой линии.
Мозжечок рыб принято подразделять на тело, вальвулу и вестибулолатеральную долю. Последняя представляет собой наиболее каудальную часть мозжечка, которая является непосредственным продолжением акустико- латереальной области. Lobus vestibulolateralis разована плотной массой гранулярных клеток вдоль латерального края доли и auriculi -другой группой гранулярных клеток, расположенных более медиально. Развитие мозжечка связано с общими специализированными электрорецепторными образованиями на теле животного.
Средний и передний мозг. Информация от органов боковой линии поступает в структуры среднего мозга. Соответствующий центр расположен в torussemicircularis покрышки среднего мозга. Это ядро может быть подразделено на две части: pars lat. u parsmed. Nucl. Lat, которое связано с акустик – латеральной областью с помощью латерального лемнического пути, о чем свидетельствуют эксперименты с дегенерацией нервных волокон после экспериментального повреждения долей боковой линии продолговатого мозга. Латеральная система рыб широко представлена в различных отделах мозга животных.
В продолговатом мозге пластиножаберных рыб ВП локализуются в пределах акустико — латеральной области. У пластиножаберных рыб ответы регистрируются в аурикулах и задней доле тела мозжечка.
Средний Он является коллектором информации, идущей от различных сенсорных систем. У неэлектрических костистых рыб акустико- латеральная система представлена в двух ядрах среднего мозга.
Биологическое значение органов боковой линии. Боковая линия дает возможность рыбе ориентироваться, чувствовать течение воды или приближение в темноте различных предметов. Органы боковой линии расположены в канале, погруженном в кожу, который сообщается с внешней средой при помощи отверстий в чешуе. Снаружи виден ряд отверстий. Эти отверстия связаны с каналом, расположенным в коже. В канале находятся чувствительные клетки, нервные окончания соединенные с нервом, идущим под кожей. Боковая линия воспринимает направление и силу тока воды. Благодаря боковой линии даже ослепленная рыба не натыкается на препятствия и способна ловить движущуюся добычу.
Биологическое назначение механорецепторной системы боковой линии еще недостаточно выявлено. Методом выработки условных, пищевых рефлексов в сочетании с денервацией органов, удалось показать о существовании некоторых поведенческих реакций при локальном действии слабых потоков воды на голову и тело у пластиножаберных и костистых рыб, а также у амфиогии. Выявлена способность ослепленных животных воспринимать искусственные препятствия на расстоянии 0,5 км, что свидетельствует от участии органов боковой линии в ориентировочном поведении с относительно малым радиусом действия. Характерно поведение многих рыб во время нереста, когда партнеры раздражают друг друга слабыми потоками воды, возникающими при движении плавников. Отмеченные реакции наблюдаются только у животных с интактной системой боковой линии.
Одной из разновидностей механической стимуляции являются вибрационные воздействия. Частотный диапазон восприятия вибрации находится в пределах от 1- до 25 Гц. Рыбы, обитающие в поверхностных слоях воды, воспринимают вибрационные стимулы в виде волн, распространяющихся по поверхности воды. В боковой линии этих рыб имеются невромасты не только вдоль боковой поверхности, но и на нижней части туловища. Такая система организации невромастов, различно расположенных на теле рыб, способствует как дифференцированному восприятию внешних стимулов, так и появлению большей чувствительности и точности локализации источников раздражении.
Относительно роли органов боковой линии в формировании стайного поведения рыб сведения противоречивы. По данным Дислера, становление стайности наблюдается у молоди карповых и окуневых рыб в конце личиночного начало малькового периода, когда органы боковой линии уже хорошо развиты. Очень интересно устроена у рыбы так называемая боковая линия, ее можно заметить почти у всех рыб. Расположена она с боков на чешуе в виде черных точек (это отверстия в чешуе) и тянется от головы до хвоста. У некоторых видов рыб например у щуки, этот орган находится и на голове, на верхней и нижней челюстях.
Боковая линия—это своеобразная, очень чувствительная сигнальная установка, ею рыба чувствует направление течения воды и его силу, ею же по малейшим изменениям тока воды может чувствовать близость препятствий на своем пути: берега, камни, коряги, завалы, движущиеся в воде предметы, проплывающую рыбу. При помощи боковой линии хищные рыбы получают сигнал о приближении живой добычи и месте ее нахождения, а “добыча” получает в это же время противоположный сигнал надвигающейся опасности.
Боковая линия особенно хорошо развита у щуки, трудно установить, каким органом — зрением или боковой линией щука более руководствуется при поисках корма. Это необходимо учесть рыболовам-спиннингистам и изготовлять свои приманки (насадки), блесны, рассчитывая не только на восприимчивость их зрением щуки, но и колебаниями блесны, раздражителем боковой линии, и чем эти колебания будут более сходны с колебаниями от живой, плывущей рыбки, тем более вероятной будет хватка щуки.
Чувствительность бокового органа очень велика, и рыба способна воспринимать очень незначительные колебания. Интересно, что рыба очень точно находит место, от которого исходит раздражение, и безошибочно захватывает предмет, даже если он приближается с заднего конца тела рыбы. Ослепленная щука могла хорошо захватывать двигающуюся около нее рыбу, однако, если рядом с ней клалась мертвая рыба, то щука ее не захватывала. Если же мертвую рыбу двигали около щуки, она брала ее как живую. Если повредить боковой орган, то рыба в темноте будет натыкаться на предметы, находящиеся в воде.
Последними научными данными установлены многие ранее неизвестные явления в жизни рыбы, например, рыбу удается приучить к тому, что она сможет отличать квадрат от прямоугольника, овал от круга и т. п. или брать корм лишь из кормушки, окрашенной в красный цвет. Рыба (пескарь) скоро научилась добывать корм именно из кормушки красного цвета, не обращая внимания на кормушки других цветов. Этим было точно установлено, что рыба различает форму и цвета. Однако это не значит, что рыба, различая цвета, находящиеся в неподвижном состоянии, сможет различить цвета быстро движущегося предмета в воде, например цвет “Девона” — белый он или желтый.
Предполагается, что боковой линией воспринимаются низкочастотные звуковые волны от 5 до 600 герц. Рыбы способны воспринимать весь диапазон звуковых колебаний: от инфра — до ультразвуковых. Установлено, что рыбы способны уловить в 10 раз меньшее изменение частот, нежели человек, но музыкальный слух у рыб в 10 раз хуже.
Раздражителями для чувствительных клеток боковой линии являются токи воды, вибрация и перепады давления, при этом происходит движение жидкости в каналах, приводящее к смещению волосков чувствительных клеток. Это воспринимается латеральными нервами туловища и головы.
С помощью органов боковой линии рыба ориентируется в потоке воды, воспринимает движущиеся предметы, различает низкочастотные колебания. Парные органы, находящиеся в боковой линии рыб, панорамно воспринимают звуковые колебания, что дает возможность рыбам четко устанавливать направление и место источника колебания. Рыбы выделяют ближнюю и дальнюю зоны акустического поля. В ближней зоне они четко определяют местонахождение источника колебания.
Инфразвуки или вибрации — колебания с частотой до 16 Гц — не воспринимаются рыбами во взвешенном состоянии без контакта с другими предметами и грунтом. Механические раздражения, возникающие в результате гидродинамических процессов, воспринимаются органами боковой линии. Звуковые колебания воспринимаются лабиринтовым аппаратом внутреннего уха. Низкочастотные звуковые колебания воспринимаются рецепторами плавательного пузыря и боковой линии.
Рецепторы боковой линии воспринимают звуки на близком расстоянии, с помощью внутреннего уха происходит анализ звуков, доносящихся с большого расстояния.
Рыбы могут определить и направление источника звука, но точно локализовать его не могут. Правильную локализацию источника звука они могут; определить только на небольшом расстоянии с помощью боковой линии. Рыбы I могут выделять нужные звуковые сигналы из шумов. Особенно хорошо они различают однотонные звуки, которые подаются ритмично. Центр, регулирующий работу акустико-латеральной системы, расположен в продолговатом мозге, он объединяет работу ядер VII, VIII и X пар нервов.
Электрорецепторная система. Способность животных находить направление своих перемещений во время миграции хорошо известна. Еще в прошлом веке высказывались предположения, что мигрирующие животные могут использовать для целей ориентации и навигации электрические и магнитные поля.
Многие слабоэлектрические рыбы обитают в мутной воде тропических рек, где видимость крайне мала, и они обладают вследствие этого слабо развитым зрением. В условиях ограниченного видения использования для ориентации в пространстве активной электросенсорной системы, включающей в себе электрический орган и систему электрорецепторов. Хорошо развитые у слабо электрических рыб органы слуха, осязания и обычная система боковой линии, также используются в процессе ориентации. Потенциалы достигают 100 мкВ/ см, а их частотных компоненты находятся в диапазоне 0-10 Гц. Функцианальные возможности электрорецепторов перекрывают диапозон электрических полей, создаваемые гидробионтами в естественных условиях.
§
Поведение рыб развивается в ходе — онтогенеза, постепенно усложняясь и становясь все более разнообразным. Самой простой двигательной реакцией является к и не з, т. е. увеличение двигательной активности в ответ на неблагоприятные или раздражающие воздействия. Кинезы наблюдаются уже на поздних стадиях эмбрионального развития.
Движение эмбриона внутри яйцевой оболочки учащается в ответ на понижение содержания кислорода, что способствует улучшению газообмена, а также более быстрому разрыву оболочки и выклеву. Вызвать резкие движения эмбриона в икринке можно воздействием раздражающих веществ, например уксуса, спирта. Кинез способствует перемещению личинок из мест с плохими условиями обитания в лучшие условия, где подвижность личинок уменьшается. Кинез не имеет направленности, но приводит к перемещению в лучшие места. Характер кинеза носит так называемая паническая оборонительная реакция таких мелких стайных рыб, как верховки, атерины, песчанки. Их быстрое беспорядочное движение мешает нападающему хищнику сосредоточить погоню за отдельной жертвой.
Более сложной формой поведения является т а к с и с или направленное перемещение. Таксис может быть отрицательным (избегание) и положительным (привлечение). Личинки лососей имеют отрицательный фототаксис — днем они прячутся в гравии, что спасает их от хищников. Положительный фототаксис личинок карповых рыб помогает им избегать заморов вблизи дна. Отношение к условиям среды и направление таксисов могут меняться с возрастом, в результате метаморфоза, в ходе сезона и по многим другим причинам.
Пестрятки семги — типичные донные одиночные оседлые рыбы. Они тяготеют к дну, камням и другому субстрату, охраняют занятую территорию от особей своего вида, имеют некоторую светобоязнь, стремятся при преследовании плыть против течения, при испуге затаиваются, легко меняют окраску под цвет окружающего грунта. После смолтификации — серебрения и подготовки к скату в море — они собираются в стаи, теряют взаимную агрессивность, при преследовании быстро уплывают за пределы видимости, держатся у поверхности и в толще воды и при испуге скатываются вниз по течению.
Подобные изменения в поведении наблюдались на японской рыбке аю. (рис.83). Половое и родительское поведение появляется у рыб по достижении половой зрелости и при инъекции некоторых гормонов, например, пролактина.
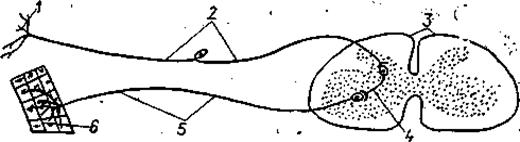
Рис. 81 . Рефлекторная дуга:
1- кожный рецептор; 2 -афферентное волокно; 3 — спинной мозг;
§
Вставочный нейрон; 5—эфферентное волокно; 6 — мышца
В формировании различных компонентов «поведенческих» реакций имеют значение, как генетические факторы, так и различные влияния внешней среды.
Кора головного мозга, имеющая большое значение в выработке условных рефлексов у высших животных, у рыб отсутствует. Однако рыбы обладают способностью вырабатывать условные рефлексы. Эти работы были начаты в лаборатории И. П. Павлова его сотрудником Ю. П. Фроловым. Исследования проводились на треске. Решено было выработать условный рефлекс на звук. В воду аквариума, где находилась треска, был опущен в запаянной коробке телефон (условным раздражителем являлся звук). Включали телефон, через 2 —3 с. включали индукционный ток (безусловный раздражитель), на что рыба реагировала движениями тела. Через некоторое количество подкреплений реакции электрическим током рыба, не дожидаясь действия электрического тока, реагировала на звук телефона сначала слабыми, а затем и более сильными движениями, как бы предупреждая действие электрического тока. Ввиду того, что рыба могла воспринимать звуки органом боковой линии, а не с помощью слуховых рецепторов, в дальнейшем телефон подвешивали над аквариумом. Результат был тот же. Этими опытами была доказана, во-первых, возможность образования условных рефлексов у рыб, во-вторых, наличие слуха.
Условные рефлексы у рыб имеют некоторые особенности, связанные с отсутствием коры голового мозга, и вырабатываются они у рыб с трудом. Они, отличаются нестойкостью, трудностью выработки дифференцировок. У рыб отсутствует высокоразвитая дифференцировочная способность к многочисленным агентам внешней среды. Они различают ограниченное количество раздражателей, которые характеризуют основные, но простые свойства среды обитания.
Рис.82. Типы поведения рыбок Аю (а) и вид с моста (б) на расположение охраняемых участков:
1 — движущаяся стая; 2 — кормящаяся стая; 3 — кормящаяся одиночка; 4 — владелец участка; 5 — движущаяся одиночка
У миног, ганоидных и поперечноротых рыб условные рефлексы отличаются нестойкостью в течение одного опыта, они быстро ослабевают и угасают на другой день. У костистых рыб условные рефлексы вырабатываются быстрее, регулярно проявляются как в течение опыта, так и на следующие дни. В. Л. Бианки и А. М. Винницкий вырабатывали — у рыб дифференцировку на величину освещенного круга, на различные фигуры (квадрат и треугольник, квадрат и прямоугольник), различные буквы (К и Б).
Большое практическое значение может иметь выработка оборонительных условных рефлексов на хищника у молоди, выпускаемой рыбоводными заводами в естественные водоемы. Молодь, выращенная на заводе, легко становится добычей хищников, так как не имеет ни индивидуального, ни зрелищного опыта общения с врагами.
В условиях рыбоводных предприятий рыбы быстро привыкают к определенному времени кормления, виду корма, способу раздачи корма. Они демонстрируют довольно точное чувство времени и временную память. Это — тоже выработка условных рефлексов.
В демонстрационном аквариуме «Абурацубо» (Япония) посетителей развлекают показом несложной дрессировки рыб: «Вперед за «знаменем»; «Спасайся, кто может!», «Плавание сквозь обруч», «Рыбья консерватория», «Рыбья арифметика» и др. Нередко наблюдается самодреосировка рыб. Если у пруда или бассейна поместить кормушку, роняющую порцию корма в ответ на нажатие рычага, дерганье за бусинку и другие устройства, рыбы осваивают это устройство в течение нескольких суток.
Личинок рыб можно успешно обучать охоте за кормовыми беспозвоночными. Если первые попытки бывают положительны на 10%, то через несколько суток уже на 70—90%.
На Конаковском рыбоводном заводе крупные карпы-производители просят корм у обслуживающего персонала и случайных посетителей, выставив из воды разинутые усатые рты.
Используя условные рефлексы исследуют различные стороны биологии рыб как спектральную чувствительность глаза, способность различать силуэты, силу и частотные характеристики звука, пороги вкусовой чувствительности, роль различных отделов нервной системы, действие наркотиков и токсикантов.
В естественной среде поведение рыб — зависит от их образа жизни. Стайные рыбы обладают способностью к согласованным маневрам. Появление хищника или кормовых организмов у одного края стаи заставляет соответствующим образом реагировать всю стаю, включая особей, которые не могли видеть раздражитель. В данном случае они демонстрируют подражание.
Одной из разновидностей подражания является движение за лидером — особью, в поведении которой не видно элементов колебаний. Часто в качестве лидеров выступают особи, имеющие индивидуальный опыт в данной ситуации. Например, карпы легче обучаются брать корм на лету, если к ним подсажена форель. Дикие карпы в аквариуме быстрее начинают брать корм, если они посажены к «ученым». Иногда лидером может служить ненормальная особь, например рыба с разрушенным передним мозгом я отличающаяся особо «прямолинейным поведением».
Стайные пелагические рыбы, как правило, равнозначны. Донные рыбы чаще бывают, оседлыми, охраняют территорию, обеспечивающую им пропитание, убежище, нерестилище.
При групповом обитании рыб может возникать «социальная иерархическая организация» с доминирующими и подчиненными рыбами. Например, в аквариуме с самцами мозамбикских тиляпий главным является самый темный по окраске, следующий в иерархии — светлее, а подчиненные самцы по окраске не отличаются от самок и в нересте не участвуют. У некоторых видов морских окуней подчиненные самцы могут превращаться в самок, доминирующие самки могут маскулинизироваться- превращаться в самцов под действием собственной гормональной системы.
Половое поведение рыб может достигать значительной сложности, включая элементы «демонстрации красоты», «ухаживания», «соперничества». Ряд рыб строят «гнезда» — роют грунт, очищают камни и травинки, сооружают пенистые шапки, даже склеивают своей слюной настоящие сооружения из водорослей.
Сложное нерестовое и родительское поведение характерно для рыб с невысокой индивидуальной плодовитостью. Некоторые рыбы заботятся об икре, личинках и даже мальках — поднимают упавших, носят их ворту. Молодь некоторых видов пасется вблизи родителей, прячется у них в ротовой и жаберной полостях. Мальки некоторых рыб живут под парусом или между щупальцами жгучих медуз. Пластичность или изменчивость поведения рыб велика. Они могут существовать в условиях, имеющих мало общего с их естественным обитанием.
Основным фактором, которые влияют на поведение рыб — Вселенная, Солнце и другие космические объекты и явления. Все живое на Земле является неотъемлемой частью Космоса, мы «впаяны» в межзвездное вещество, 97% которого, состоит из мельчайших частиц – нейтрино, наше тело постоянно пронизывают миллиарды этих частиц. Земля и Космос – единая колебательная система, и мы связаны друг с другом с другом невидимыми силами. Нa один квадратный метр земной поверхности ежесекундно падает более 10000 заряженных космических частиц, которые составляют космическое излучение, и есть теория, согласно которой это излучение руководит земной погодой.
Контрольные вопросы.
1.Какие функциональные группы нейронов имеется в ЦНС?
2.Какую роль выполняют группы нейронов? Как осуществляется взаимодействие нейронов?
3.Каковы особенности строения и функционирования межнейронных синапсов?
4.Что такое торможение и какие виды имеют место у рыб?
5.Каково строение и функциональное назначение рефлекторной дуги?
6.Изобразите рефлекторную дугу и выделите основные звения дуг.
7.Изобразите на схеме расположение разных отделов головного мозга, перечислите входящие в их состав структуры.Опишите строение спинного мозга.
8. Какие центры регуляции расположены в продолговатом мозгу? Какое отношение имеет продолговатый мозг к двигательной функции рыб?
9.Укажите основные функции мозжечка.
10. Какие структуры среднего мозга и как осуществляет регуляцию тонуса мышц в зависимости от положения тела в пространстве?
11.Как вы понимаете проводниковую и рефлекторную функции спинного мозга?
12.Что такое поведение рыб и какие физиологические механизмы лежат в его основе? Охарактеризуйте врожденные нормы поведения рыб и как они регулируюся? Дайте характеристику условных и безусловных рефлексов Что такое высшая нервная деятельность?
Глава 12
Любой фактор, воздействующий на живое существо, природа ответила созданием у этого существа воспринимающего органа чувств. Каждый организм способен существовать и выживать только в том случае, если он имеет возможность получать информацию о всех действующих на него внешних факторах и приспосабливаться к существованию в этой переменчивой обстановке.
Информацию об окружающей среде организм получает с помощью системы органов чувств. Исследователями обнаружено, что рыбы обладают целым набором органов чувств, число которых намного превосходит упомянутые в существующей ныне классификации, все существа имеют анализаторы, способные определять физические поля, в том числе электромагнитные, магнитные, электрические, гравитационные, и что они обладают барочувствительностью и способностью воспринимать геомагнитные поля. С помощью этих анализаторов живые существа принимают метеорологическую информацию, помогающую им предсказывать погоду, осуществляют локацию, определяют биологическое время по плоскостям поляризованного света или же с помощью улавливания космического излучения, магнитного поля Земли и учета самодвижения в космическом пространстве Солнца, Земли и Луны. С помощью хеморецепторов точно ощущают состав примесей в воде, используют почти все известные оптические приспособления, осуществляют объемное звуковосприятие и звуковое кодирование предметов, обмениваются между собой информацией, определяют конфигурацию молекул и т.п. видимо, это только часть возможностей органов чувств рыб.
Официально было объявлено об открытии у рыб «шестого» чувства — сейсмического слуха. Под шестым органом чувств, подразумевали в одном случае боковую линию рыб, в другом орган, отвечающий за электролокацию.
По литературным данным, в настоящее время говориться о 19 органах чувств у рыб. Оказывается, рыбы не отстали от нас в своем развитии на сотни миллионов лет, как считали до сих пор, а превзошли людей и всех наземных животных в способности получать извне информацию о внешних воздействиях. Если признать наличие этих органов, то откроется более широкий путь решения многих проблем, связанных с поведением рыб.
Имеющаяся классификация органов чувств рыб, составленная с учетом восприятия ими внешних воздействий, состоит из шести групп: механорецепторов, хеморецепторов, терморецепторов, а также рецепторов звуковых, световых и болевых. В настоящее время, к имеющимся рецепторам надо добавить еще пять групп органов чувств: электрорецепторы, магниторецепторы, гравитационные рецепторы, барорецепторы, виброрецепторы.
Таким образом, новая рабочая классификация состоит из следующих органов чувств: зрения, световой чувствительности, слуха, чувствительности, обоняния, осязания, вкуса, локационной, холодовой, тепловой, гравитационной, равновесия, барочувствительности, болевой, вибрационной, электролокации, электросейсмочувствительности, электроосязания, магнитной чувствительности, магнитно-азимутной ориентации.
Жизнь рыбы становится возможной благодаря наличию адекватных реакций ее организма на изменения, происходящие во внешней среде или в самом организме. Получение информации о физических и химических характеристиках среды называется
рецепцией, труктуры организма, воспринимающие эту информацию, называются рецепторами – приемниками или анализаторами.
Рыба реагирует на световое излучение и внешний вид объектов материального мира: обладает зрением, реагирует на температуру окружающей среды – терморецепцией. На поведение рыбы влияют многие растворенные в воде вещества. Рыба обладает химической рецепцией — вкусом и обонянием, воспринимает упругие колебания среды — слышит. Рыба способна к проприорецепции — анализу своей позы и положения тела, кроме того, она способна реагировать на ускорения, вызываемые смещением тела. Легко обнаруживается тактильная рецепция рыб — реакция на прикосновения, удар, давление. На рыб оказывает влияние электрический ток, а также в некоторых условиях она может реагировать на магнитное поле. На радиоактивное излучение сублетальной интенсивности рыбы, как и другие живые существа, не реагируют.
12.1 Органы зрения и механизм восприятия изображения.
Как у большинства живых существ органом зрения у рыб являются глаза. Анатомически и функционально глаза рыб не сильно отличаются от глаз других животных. Работа глаз у всех животных основана на едином принципе: свет действует на светочувствительные субстанции, разлагая их так же, как это происходит с соединениями серебра при обработке фотопленки и фотобумаги для получения на них изображения. Но фотопроцессы необратимы, т.е. соединения серебра не восстанавливаются в отличие от родопсина и йодопсина, которые являются светочувствительными субстанциями палочек и колбочек — рецепторной основой сетчатки глаза.
Рыбы живут не в атмосфере, а в полупрозрачной среде, где видимость (даже в самой прозрачной воде) не превышает 40 м. В большинстве же водоемов видимость составляет всего несколько метров, а есть водоемы, где она равна нулю.
Орган зрения рыб является самым несовершенным из всех органов чувств, которыми они располагают. Но несовершенными являются не все функции глаз рыб. К функциям глаз относятся контрастная чувствительность, острота зрения, быстрота различения деталей и устойчивость ясного видения. Контрастная чувствительность — очень важная функция. Она заключается в способности глаз воспринимать границу яркостей двух смежных предметов, видеть их раздельно существующими. Яркость — это сила света, отраженного от поверхности предмета, она является единственной световой характеристикой предмета, который видит глаз.
Контрастная чувствительность глаза повышается при увеличении яркости света, но до определенного уровня, что объясняется слепящим действием света. Острота зрения также связана, со способностью глаза видеть раздельно две детали предмета. Быстрота различения деталей увеличивается при увеличении освещенности, но также до известных пределов. Способность сохранять в течение некоторого времени в поле зрения фиксированные глазом детали называют устойчивостью ясного видения, она также зависит от условий освещения. Для нормального видения рыбам необходимы следующие условия: достаточная и равномерная освещенность предметов, постоянство освещенности во времени, благоприятное распределение яркости в поле зрения, отсутствие прямой и отраженной блескости.
Работа зрительного анализатора основана на определении цвета предмета, его формы, передвижения, размеров, удаленности и расположения по отношению к другим. Чувствительность глаза может меняться в зависимости от освещенности в 200000 -250000 раз, а пороговая чувствительность определяется энергией, равной всего лишь нескольким квантам.
Рыбы обладают панорамным зрением, они видят в нескольких направлениях одновременно. Контрастная чувствительность у рыб глаз гораздо выше, чем у человека, но острота зрения в 2-3 раза ниже (Рис.83).
Глаза у рыбы хорошо развиты и расположены у большинства по бокам головы. Видит рыба ими на близком расстоянии превосходно и не только в воде, но и вне воды.
Угол зрения рыбы (включая движения глаза) по горизонтали достигает 320—340° и по вертикали около 300°, так что рыба даже при неподвижном состоянии тела способна видеть большую часть окружающей среды (рис. 83,1).
 Как рыбы видят предметы, находящиеся вне сферы воды, можно понять, принимая во внимание закон преломления световых лучей. Это станет ясным, если мы посмотрим на рисунок, представляющий пруд, наполненный водой (рис.83-2,3).
Как рыбы видят предметы, находящиеся вне сферы воды, можно понять, принимая во внимание закон преломления световых лучей. Это станет ясным, если мы посмотрим на рисунок, представляющий пруд, наполненный водой (рис.83-2,3).
Если бы пруд был пуст, то из точки «О» можно было бы видеть только те предметы, которые находятся в пространстве между ОА и 0В. Другое дело, если пруд наполнен водой, тогда рыба может из точки «О» видеть предмет «И», так как световой луч вследствие перехода из воды в воздух преломляется в точке «М» и вследствие этого край пруда не заслоняет от рыбы предметов, находящихся на некотором расстоянии от берега. Другими словами, пространство видимости для рыб шире, чем мы думаем, и это небесполезно запомнить всем рыбакам, как любителям, так и профессионалам. Если поэтому к точке «М» приблизится человек, то рыба со дна пруда увидит его ранее, чем он успеет подойти к берегу, стоящего на берегу пруда рыба видит с головы до ног, а не только верхнюю часть туловища.
Показатель преломления воды равен 1,33. Луч, падающий перпендикулярно к поверхности, при переходе из воды в воздух не будет преломляться. При увеличении угла падения будет увеличиваться и угол преломления. Когда же угол преломления достигнет 90°, луч не выйдет из воды, а будет проходить вдоль ее поверхности. При дальнейшем увеличении угла падения свет совсем не будет выходить из воды, полностью отражаясь от поверхности.
Рыбы близоруки, некоторые из них четко видят предметы только на расстоянии 5 см от глаз. Самые «зоркие» рыбы в состоянии различать предметы на расстоянии 10-15 м, а четко их видеть — только в пределах 1м. Рыбы воспринимают оттенки цветов спектра в 10-20 раз хуже, чем человек. Область предпочтительных цветов у рыб такая же, как у человека — голубые и светло-зеленые тона. Многие костистые рыбы обладают цветным зрением. Скаты цвета не различают, т.е. являются ахроматами. У рыб, как и у человека, имеется три вида колбочек, воспринимающих три разных цвета. Одни колбочки реагируют на насыщенный красный цвет, другие на насыщенный зеленый, третьи — на насыщенный сине-фиолетовый цвет. В зависимости от количества и степени возбуждения колбочек воспринимаются разные оттенки цветов.
Из наблюдений в аквариуме, в экспериментальных условиях рыбы собирались в желто-зеленой и зеленой частях спектра, установлено, что особенно хорошо рыбы отличают красный цвет. При разведении рыб нетрудно заметить, что для нереста большую роль играет цвет субстрата. Так, при употреблении искусственных субстратов вместо водных растений предпочтение отдается окрашеным в зеленый цвет и коричневатый. Характер окраски играет определенную роль и у стайных рыб при их соединении в стаю.
Разные рыбы по-разному реагируют на цвета. Отмечают, что для рыб, живущих в прозрачной воде, оптическая сигнализация в жизни имеет большее значение, чем для рыб, живущих в непрозрачной среде. Зрение имеет наибольшее значение для рыб, питающихся планктоном. Хищные рыбы при преследовании жертвы пользуются зрением только для корректировки движения перед броском на добычу.
Многих рыб привлекают зрительные ориентиры — подвижные и неподвижные предметы, находящиеся на пути их движения. Замечено, что в большей степени рыб привлекают темные предметы. Не исключено, что эти предпочтения рыб связаны с обстоятельствами их миграционной деятельности.
Рыбы реагируют и на искусственный свет. Он действует на них по-разному, одних привлекает, других отпугивает. Считается, что к искусственному источнику света идут рыбы, которые в поисках пищи ориентируются на зрение.
В лунные ночи активность отдельных видов рыб повышается. Хищники легче находят добычу. Некоторые рыбы (особенно язь) при сильном лунном свете и даже при искусственном освещении уходит на более глубокие или затененные места подводного мира. Рыбы, не выносят сильного освещения, в этом случае они не способны различать слабоосвещенные предметы. К тому же при очень сильном свете не восстанавливается родопсин глаз — рыбы слепнут
Глаз у рыб является главным органом зрения, хотя имеются и некоторые другие светочувствительные структуры, имеющие различную степень развития у разных видов. У миног в области хвоста имеются светочувствительные клетки. К восприятию света способна железа внутренней секреции — эпифиз. Развитие зрения зависит от той роли, которую оно играет в жизни данного вида рыб. У пещерных рыб, миксин, личинок миног глаза недоразвиты. У этих рыб хорошо развиты другие органы чувств — вкуса, осязания, орган боковой линии. У рыб, живущих в условиях хорошей освещенности (щука, окунь), глаза развиты хорошо, а у придонных видов (вьюн, сом), они развиты хуже.
Некоторые темнолюбивые виды (лещ, ерш, густера) имеют хорошо развитые глаза. Рыбы, живущие на больших глубинах, куда еще проникают лучи света, имеют огромные глаза, улавливающие слабый свет, идущий сверху, и свечение, исходящее от живых организмов. У глубоководных рыб могут быть большие глаза, но часто они недоразвиты.
В зависимости от расположения глаз различают рыб с монокулярным зрением, поля зрения двух глаз не перекрещиваются и рыб с бинокулярным зрением, когда поля зрения двух глаз перекрещиваются. У большинства рыб глаза двигаются согласованно. У камбалы, морских игл и некоторых других рыбглаза могут двигаться независимо друг от друга.
Своеобразное строение глаз имеет четырехглазка, живущая в лагунах у берегов Центральной и Южной Америки. Она плавает в поверхностных водах таким образом, что половина ее головы находится на воздухе, а вторая половина — в воде. Соответственно и глаза разделены горизонтальной эпителиальной перегородкой на две части — верхнюю и нижнюю. Верхние части глаз имеют плоский хрусталик, как у наземных животных, а нижние — округлый, как у водных животных.
Строение глаза рыб имеют некоторые особенности, связанные с жизнью в водной среде. У рыб нет век, глаза их постоянно открыты. Лишь у некоторых акул имеется, прозрачная мигательная перепонка, закрывающая глаз с внутреннего угла. У кефали и некоторых сельдей имеется жировое веко, частично закрывающее глаз. У рыб отсутствуют слезные железы, так как в отличие от наземных животных им нет необходимости смачивать глазное яблоко.
Различия в величине глаз, определяются влиянием двух комплексов факторов в первых, той ролью, которую играет зрительный рецептор в общей системе рецепторов органов чувств рыбы, во-вторых, общим характерно морфологической организации рыбы. Величина глаз у рыб варьирует в широких пределах, громадные, телоскопические; маленькие, или даже атрофированные, полностью скрытые под кожей и не функционирующие.
Уменьшение размера глаз, их атрофии, наблюдается, когда освещенность уменьшается, некоторые случаях это приводит к обратному эффекту — появлению телескопических глаз, при помощи которых организации получает возможность улавливать очень слабый свет.
При хорошей освещенности сами глаза развиты нормально, а размер глаз относительно уменьшается, атрофия глаз характерна, для глубоководных рыб. Атрофия глаз наблюдается также у рыб пещер и подземных источников. У придонных рыб, зрения бывает слабой или ноль, в результате чего размер глаз значительно уменьшается. Атрофия глаз, компенсируется усиления прочих органов чувств, особенно органов осязания, вкуса боковой линии. Развитые чувства осязания и вкуса, выражается наличием специальных подвижных придатков – усов на которых расположены соответствующие рецепторы.
Горизонтальный диаметр глаз составляет 3-10% длины тела до конца позвоночного столба. Увеличение размеров глаз наблюдается у глубоководных форм. У мелких форм, глаза крупные, а у крупных мельче.
Величина глаз зависит также от относительной высоты тела отмечено, что телоскопические глаза имеются обычно у видов с массивным, высоким телом. Поскольку глаза, как и все остальные, части тела, и внесены в единый ансамбль внешней организации рыбы, естественно, что с уменьшением высоты тела величина вертикального диаметра глаза все более жестко лимитируется.
С уменьшением относительно высоты тела размеры глаза прогрессивно уменьшается. У рыб в онтогенезе относительная величина глаза первоначального увеличивается, а затем уменьшается. На ранних этапах развития, величина глаза лимитируется высотой тела и увеличивается по мере увеличения высоты тела личинки. Расположение глаз у рыб определяется особенностями их экологии. У пелагических и природно- пелагических видов планктоноядных, хищных или растительноядных – глаза располагаются обычно по бокам головы примерно на уровне продольной оси тела. В этом случае глаза находятся в одинаковом положении по отношению как к верхней полусфере пространства, так и нижней. У видов природных, независимо, от характера их питания, глаза находятся на верхней стороне головы.
Строение глаза рыбы изображено на рис.84. Хрусталик шарообразной формы, не изменяет формы и почти касается плоской роговицы, поэтому рыбы близоруки, век нет. Зрачок не может сужаться и расширяться. Благодаря сокращению мышц серповидного отростка хрусталик глаза может оттягиваться назад, поэтому, большинство их различают предметы на расстоянии до 1 м, а максимально видят не более чем на 10-15м
Снаружи располагается склера, или белковая оболочка, которая впереди переходит в прозрачную роговицу. Склера состоит из плотной хрящевой ткани. Роговица — представлена многослойным плоским эпителием, она прозрачна, только в краевой ее зоне имеются кровеносные сосуды. Роговица глаз рыб очень слабо выпукла. За склерой расположена серебристая оболочка, состоящая из нескольких слоев плоских клеток, содержащих кристаллы гуанина. Далее находится сосудистая оболочка, содержащая сосуды и пигментные клетки. В передней части глаза она переходит в радужную оболочку, в центре которой имеется отверстие — зрачок. Через зрачок — световые лучи проникают внутрь глаза. Зрачок может расширяться и сужаться — благодаря работе особых мышц, таким образом, регулируется поступление в глаз света.
Реакция зрачка на свет хорошо выражена у акул и угрей. К. пигментному слою примыкает сетчатка. Отростки пигментных клеток могут укрывать светочувствительные клетки, так называемые палочки и колбочки. Палочки оценивают степень освещенности, силу света, колбочки осуществляют цветовосприятие. Палочки функционируют при дневном свете и в сумерках, колбочки — только при сильном дневном, освещении. Пигментный слой уменьшает рассеяние света. За слоем светочувствительных клеток следует слой биполярных нервных клеток, а за ними идет слой ганглиозных клеток, аксоны последних дают начало зрительным нервам. Там, где оптическая ось глаза пересекает сетчатку, расположено желтое пятно. Желтый цвет этой области зависит от присутствия каротиноидов. В центре желтого пятна расположено небольшое углубление — центральная ямка. На эту область сетчатки проецируются те объекты, которые находятся перед глазами и наиболее отчетливо видны. У дневных рыб здесь раеполагаются только колбочки.
По периферии центральной ямки располагаются палочки, которые обусловливают периферическое зрение. Предметы, равно как гамма цвета, проецирующиеся на эту область сетчатки, видны неотчетливо.
Палочки и колбочки состоят из внутреннего и наружного сегментов (члеников). Наружный сегмент содержит несколько сотен тончайших дисков, он соединен с внутренним сегментом с помощью ножки.
Внутренний сегменг содержит характерные для клетки органеллы. Митохондрии образуют компактную массу — эллипсоид, который прилегает к наружному сегменту. Эллипсоид соединяется тонкими нитями протоплазмы (сократительный элемент — миоид) с ядром.
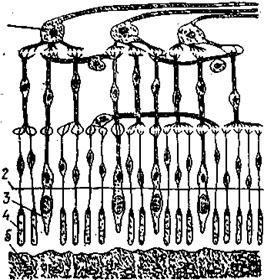
Рис.85. Строение сетчатки глаза:
1 — ганглиозные клетки;
2–наружная пограничная мембрана;
3 — колбочки;
4- палочки;
§
Палочки имеют длинные цилиндрические наружные — сегменты, у колбочек наружные сегменты короткие, каплеобразные. Наружные сегменты зрительных клеток являются их рецепторным аппаратом (рис.85)
У костистых рыб имеются колбочки трех видов: одиночные с коротким или длинным миоидом и близнецовые колбочки, для которых характерно тесное прилегание внутренних сегментов.
У рыб, амфибий и птиц в колбочках между внутренними и наружными сегментами расположена масляная капля, которая избирательно поглощает свет. Сетчатка позвоночных инвертирована (т. е. как бы: вывернута таким образом, что к наружным сегментам фоторецепторов свет доходит в последнюю очередь). В эпифизе также имеется сетчатка с фоторецепторнымш клетками, но она не инвертирована.
У светолюбивых рыб (окунь, форель, щука) светочувствительные элементы многочисленны с небольшим преобладанием палочек, над колбочками. У этих рыб каждая фоторецепторная клетка контактирует с одной биполярной клеткой, а четыре биполярные клетки — с одной ганглиозной, что обеспечивает высокую остроту зрения при сравнительно низкой световой чувствительности.
У рыб, избегающих яркого света (лещ, ерш, угорь, налим), в сетчатке в значительной степени преобладают палочки. У них с каждой биполярной клеткой связано до десяти и более фоторецепторных, а соответственно на одну ганглиозную клетку приходятся десятки и даже сотни фоторецепторных клеток. Это приводит к уменьшению остроты зрения, но увеличивается световая чувствительность. У некоторых видов морских глубоководных рыб колбочки вообще отсутствуют.
Палочки пресноводных рыб содержат пурпурный зрительньй пигмент- порфиропаин (максимум поглощения света приходится на 520—1540 нм), палочки морских рыб—родопсин (максимум поглощения света 1500 нм) и смесь родопсина с порфиропаином (максимум поглощения света 500—640 нм). Колбочки рыб содержат фиолетовый пигмент йодопсин (максимум поглощения света 682 нм) и голубой пигмент цианолсин (650 нм). Если рыбы из морской воды перемещаются в пресную, то родопсин у них меняется на порфиропсин.
Между палочками и колбочками имеются реципромные -взаимосогласованость отношения, т. е. когда функционируют палочка, то функция колбочки заторможены, и наоборот. У морских рыб светочувствительность сдвинута в коротковолновую часть спектра, а у пресноводных — в длинноволновую. В сумерки рыбы воспринимают коротковолновую, часть спектра, днем — длинноволновую.
Виды, живущие в поверхностных слоях (керчак, морской налим), воспринимают более широкий спектр лучей света (400-760 нм), чем донные рыбы (скаты), воспринимающие более узкий спектр луча света (410-640 нм). В воде с увеличением глубины усиливается поглощение красных и ультрафиолетовых, лучей, на глубине преобладают синие лучи, соответственно суживается воспринимаемый спектр луча света.
Позади зрачка располагается хрусталик. Показатель преломления зрачка у рыб равен 1,649-1,653. Показатели преломления роговицы и стекловидного тела приблизительно такие же, как у
воды -1,33. Стекловидное тело расположено позади хрусталика и способствует сохранению формы глаза. При ранениях оно вытекает, глазные оболочки спадаются и глаз перестает видеть.
Основную преломляющую функцию у рыб выполняет хрусталик, обладающая способностью к адаптации — аккомодаций Аккомодация — способность четкого видения предметов, находящихся на разных расстояниях от глаза, у млекопитающих осуществляется за счет изменения кривизны хрусталика. У рыб имеется галлеров орган (специальные мышцы), за счет которого изменяется положение хрусталика по отношению к сетчатке. Аккомодация лучше выражена у хищных рыб, чем у мирных. В спокойном состоянии глаза рыб гипермиотропичны, т. е. выражена установка «на бесконечность». Острота зрения определяется наименьшим углом между двумя точками, при котором они воспринимаются раздельно.
Механизм восприятия света. При попадании света на сетчатку в ней происходят фотохимические, химические, электрические и ретиномоторные процессы.
К фотохимическим процессам относится распад родопсина, порфиропсина и йодопсина. Скорость распада родопсина и порфиропсина выше, чем йодопсина. Колбочки возбуждаются при действии длинноволновой части спектра, а палочки — коротковолновой. Палочки примерно в 1000 раз более чувствительны к действию света, чем колбочки. При интенсивном освещении преобладают процессы распада зрительных пигментов, в темноте — восстановительные процессы. Йодопсин восстанавливается быстрее родопсина и порфиропсина. Распад и восстановление зрительных пигментов происходят в течение нескольких секунд.
В сетчатке происходят электрические явления и процессы обмена веществ — гликолиз, распад и восстановление АТФ. Электрические явления заключаются в том, что при воздействии света в сетчатке возникают электрические потенциалы. Ретиномоторные явления наблюдаются в сетчатке таких пойкилотермных животных, как рыбы и амфибии. На свету колбочки приближаются к наружной мембране, а палочки закрываются пигментными клетками, в темноте палочки подтягиваются к наружной мембране, а наружные сегменты колбочек прикрываются пигментными клетками. Движение фоторецепторных клеток происходит за счет расслабления и сокращения миоида, в пигментных клетках пигмент перемещается по отросткам (рис.86).
Рецепторы сетчатки генерируют, импульсы при существенном изменении их освещенности. При попадании на них света они дают импульс «на включение», при сильном уменьшении освещенности- «на выключение». Если освещенность не изменяется, то информация с сетчатки в мозг не поступает, и зрительные образы становятся невидимыми. Неподвижные предметы становятся видимыми потому, что движение глаз, как бы ощупывает изображение, особенно контуры предметов. Изображение на сетчатке перевернутое и уменьшенное.
Движение воспринимается рыбами в результате так называемого киноскопического эффекта — последовательного перемещения зрительных образов с одних чувствительных элементов на другие.
В результате наличия рефрактерного периода (временной потери чувствительности после рабочего цикла) разрешающая способность ограничена. Критическая частота мельканий — вспышек света или движущихся в поле зрения полос — различается у разных видов рыб по разному. Критическая частота мельканий у верховки 14 Гц, у атерины 67 Гц, а у человека 18—24 Гц.
Пространственное зрение (рис.87), т. е. свойство восприятия предметов в пространстве, обеспечивается аккомодацией и остротой зрения. Острота зрения зависит от разрешающей способности преломляющих сред и сетчатки. Аккомодация глаза у рыб происходит за счет сокращений галлерова органа. При расслаблении галлерова органа глаз становится близоруким (миотропичным).
Зрительные нервы содержат три группы афферентных нервных волокон, каждая из которых проводит раздражение только от одного вида колбочек.
Рис.88

Рис.90 Отделы головного мозга акулы
§
Любой источник звука, находящийся на субстрате, помимо излучения классических звуковых волн, распространяющихся в водной или воздушной среде, рассеивает часть энергии, в форме различного рода вибрации, распространяющихся в субстрате и по его поверхности.
Под слуховой системой мы понимаем рецепторную систему, способную воспринимать тот или иной компонент звукоизучения, локализовать и оценивать характер источника, создавал предпосылки для формирования специфических поведенческих реакции организма.
Слуховую функцию у рыб осуществляют, помимо основного органа слуха еще и боковая линия, плавательный пузырь, а также специфические нервные окончания.
Органы слуха рыб развивались в водной среде, которая проводит звук в 4 раза быстрее и на большие расстояния, чем атмосфера. Диапазон восприятия звуков у рыб существенно шире, чем у многих наземных животных и людей.
В жизни рыб слух играет очень большую роль, особенно рыб, обитающих в мутной воде. В боковой линии рыб обнаружены образования, которые регистрируют акустические и другие колебания воды.
Слуховой анализатор человека воспринимает колебания — с частотой от 16 до 20000 Гц. Звуки с частотой ниже Гц относятся к инфразвукам, а выше 20000 Гц — к ультразвукам. Наилучшее восприятие звуковых колебаний наблюдается в интервале от 1000 до 4000 Гц. Спектр звуковых частот, воспринимаемых рыбами, по сравнению с человеком значительно сокращен. Так, например карась воспринимает звуки в диапазоне 4 (31—21760 Гц, карликовый сомик —60—1600 Гц, акула 500- 2500 Гц.
Органы слуха рыб обладают способностью к адаптации факторам окружающей среды в частности, постоянному или однообразному и часто повторяющемуся шуму, например работе землечерпалки, рыба быстро привыкает и шума ее не боится. Также не ухудшает клева рыбы шум проходящего парохода, поезда и даже люди, купающиеся довольно близко к месту ловли, не распугивают рыб. Испуг у рыбы очень кратковременен. Удар блесны о воду, если он произведен без сильного шума, не только не пугает хищника, а возможно настораживает его в ожидании появления чего-либо съедобного для него. Рыба, может воспринимать отдельные звуки, если они вызывают колебания водяной среды. Благодаря плотности воды звуковые волны хорошо предаются через кости черепа и воспринимаются органами слуха рыбы. Рыбы могут слышать шаги человека, идущего по берегу, звон колокольчика, выстрел.
Анатомически, как и у всех позвоночных, основной орган слуха — ухо является парным органом и составляет единое целое с органом равновесия. Отличие заключается только в том, что у рыб нет ушных раковин и барабанных перепонок, так как они живут в другой среде. Орган слуха и лабиринт у рыб одновременно является органом равновесия, он расположен в задней части черепа, внутри хрящевой, или костной, камеры, состоит из верхнего и нижнего мешочков, в которых находятся отолиты- камешки.
Орган слуха рыб представлен только внутренним ухом и состоит из лабиринта. Внутреннее ухо стато — акустический парный орган. У хрящевых рыб состоит из перепончатого лабиринта заключенного в хрящевую слуховую капсулу — боковое расширение хрящевого черепа позади глазницы. Лабиринт, представлен тремя перепончатыми полукружными каналами и тремя отолитовыми органами — утрикулюсом, саккулюсом и лагеной (рис. 91,92,93). Лабиринт делится на две части: верхнюю, к которой относятся полукружные каналы и утрикулюс, и нижнюю часть—саккулюс и лагена. Три изогнутые трубочки полукружные каналы лежат в трех взаимно перпендикулярных плоскостях и концами открываются в преддверье или перепончатый мешок. Он подразделяется на две части верхнюю овальный мешочек и более крупную нижнюю — круглый мешочек, от которого отходит небольшой вырост — лагена.
Полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики отоконии. В полости круглого мешочка обычно находятся более крупные известковые образования отолиты состоящие из соединений кальция. Колебания, которых воспринимаются слуховым нервом. Окончания слухового нерва подходят к отдельным участкам перепончатого лабиринта, покрытым чувствующим эпителием – слуховым пятнам и слуховым гребням. Звуковые волны передаются непосредственно через ткани восприятия колебания, которые воспринимаются слуховым нервом.
Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях. Каждый полукружный, канал впадает в утрикулюс двумя концами, один из которых — расширяется в ампулу. Здесь имеются возвышения, называваемые слуховыми пятнами, или макулами, где расположены скопления чувствительных волосковых клеток. Тончайшие волоски этих клеток соединены студнеобразным веществом, образуя купулу. К волосковым клеткам подходят окончания VIII пары-черепно-мозговых нервов.
В утрикулюсе костистых рыб находится один крупный отолит. Отолиты располагаются также в лагене и саккулюсе. Отолит саккулюса используется для определения возраста рыб. Саккулюс хрящевых рыб сообщается с внешней средой посредством перепончатого выроста, у костистых рыб подобный вырост саккулюса заканчивается слепо.
Работами Динкграафа и Фриша было подтверждено, что слуховая функция зависит от нижней части лабиринта — саккулюса и лагены.
Лабиринт связан с плавательным пузырем цепью веберовских косточек (карповые, обыкновенные сомы, харациновые, гимнотовые), и рыбы способны воспринимать высокие звуковые тона. С помощью плавательного пузыря происходит трансформация звуков высокой частоты в низкочастотные колебания (смещения), которые воспринимаются рецепторными клетками. У некоторых рыб, не имеющих плавательного пузыря, эту функцию выполняют воздушные полости, связанные с внутренним ухом.
Рис.93 . Внутреннее ухо или лабиринт рыб:
а— миксины; б — акулы; в — костистые рыбы;
1 — задняя криста; 2-криста горизонтального канала; 3- передняя криста;
4-эндолимфатический проток; 5- макула саккулюса, 6- макула утрикулюса; 7 — макула лагены; 8 — общая ножка полукружных каналов
Рыбы обладают также удивительным «прибором» — анализатором сигналов. Благодаря этому органу рыбы из всего хаоса окружающих их звуков и колебательных проявлений способны выделить нужные и важные для них сигналы, даже такие слабые, которые находятся на стадии возникновения или на грани затухания.

Рыбы способны усиливать эти слабые сигналы и затем воспринимать их анализирующими образованиями.
Плавательный пузырь, как полагают, играет роль резонатора и преобразователя звуковых волн, который увеличивает остроту слуха. Он выполняет также звукообразовательную функцию. Рыбы широко пользуются звуковой сигнализацией, они способны и воспринимать, и издавать звуки в широком диапазоне частот. Рыбами хорошо воспринимаются инфразвуковые колебания. Частоты равные 4-6 герцам действуют губительно на живые организмы, так как эти колебания входят в резонанс с колебаниями самого тела или отдельных органов и разрушают их. Не исключено, что рыбы реагируют на приближение ненастной погоды благодаря восприятию низкочастотных акустических колебаний, исходящих от надвигающихся циклонов.
Рыбы способны «предсказывать» изменения погоды задолго до их наступления, рыбы эти изменения фиксируют по разнице силы звуков, а возможно, и по уровню помех для прохождения волн определенного диапазона.
12.3Механизм равновесия тела у рыб. У костистых рыб утрикулюс является основным рецептором положения тела. Отолиты с помощью студенистой массы соединены с волосками чувствительного эпителия. При положении головы теменем вверх отолиты давят на волоски, при положении теменем вниз висят на волосках, при боковых положениях головы отмечается разная степень натяжения волосков. С помощью отолитов рыба принимает правильное положение головы (теменем вверх), следовательно, и туловища (спиной вверх). Для сохранения правильного положения тела также имеет значение информация, поступающая от зрительных анализаторов.
Фриш установил, что при удалении верхней части лабиринта (утрикулюса и полукружных каналов) у гольянов нарушается равновесие, рыбы ложатся на бок, брюшко или на спину на дне аквариума. При плавании они также принимают разное положение тела. Зрячие рыбы быстро восстанавливают правильное положение, а слепые равновесие восстановить не могут. Таким образом, полукружные каналы имеют большое значение в поддержании равновесия, кроме того, с помощью этих каналов происходит восприятие изменения скорости движения или вращения.
В начале движения или при его ускорении эндолимфа несколько отстает от движения головы и волоски чувствительных клеток отклоняются в сторону, противоположную, движению. При этом раздражаются окончания вестибулярного нерва. При остановке или замедлении движения эндолимфа полукружных каналов по инерции продолжает двигаться и отклоняет волоски чувствительных клеток по ходу движения.
Изучение функционального значения различных отделов лабиринта для восприятия звуковых колебаний проводилось с помощью исследования поведения рыб на основе выработки условных рефлексов, а также с помощью электрофизиологических методов.
В 1910 г. Пипер обнаружил появление токов действия при раздражении нижней частей лабиринта — саккулюса свежеубитой рыбы и отсутствие таковых при раздражении утрикулюса и полукружных каналов.
Позднее Фролов экспериментально подтвердил восприятие звуковых колебаний рыбами, проводя эксперименты на треске, применяя условно-рефлекторную методику. Фриш вырабатывал условные рефлексы на свист у карликовых сомиков. Штеттеэ. у сомов, гольянов и гольцов вырабатывал условные рефлексы на определенные звуки, подкрепляя их дачей мясных крошек, а также вызывал торможение пищевой реакции на другие звуки, ударяя рыбу стеклянной палочкой.
Органы локационной чувствительности рыб. Способности рыб к эхолокации, осуществляется не органами слуха, а самостоятельным органом — локационного органа чувств. Эхолокация относитсяковторому типу слуха. В боковой линии рыб имеется радар и сонар — составляющие органа локации.
Рыбы для своей жизнедеятельности используют электролокацию, эхолокацию, даже термолокацию. Электролокацию, часто называют шестым органом чувств рыб. Элекролокация хорошо развита у дельфинов и летучих мышей. Эти животные используют ультразвуковые импульсы частотой 60000-100000 герц, длительность посылаемого сигнала равна 0,0001 секунды, интервал между импульсами составляет 0,02 секунды. Это время требуется для анализа головным мозгом полученной информации и формирование специфической ответной реакции организма. У рыб это время немного короче. При электролокации, где скорость посылаемого сигнала равна 300000 км/с, времени для анализа отраженного сигнала у животного нет, посланный сигнал отразится и будет воспринят практически в одно и то же время.
Пресноводные рыбы не могут использовать для локации ультразвук. Для этого рыбы должны постоянно двигатся, а рыбам значительное время необходимо отдыхать. Дельфины же круглые сутки находятся в движении, у них попеременно отдыхает то левая, то правая половина мозга. Рыбы для локации используют низкочастотные волны широкого диапазона. Считается, что эти волны служат рыбам для коммуникационных целей.
Гидроакустические исследования показали, что рыбы слишком «болтливы» для неразумного существа, слишком много звуков они производят, притом «разговоры» ведутся на частотах, находящихся за пределами нормального восприятия их основным органом слуха, т.е. их сигналы более целесообразны в качестве посылаемых радарами рыб локационных сигналов. Низкочастотные волны плохо отражаются от мелких предметов, меньше поглощаются водой, слышны на большие расстояния, распространяются равномерно во все стороны от источника звука, их использование для локации дает рыбам возможность панорамного «видения — слышания» окружающего пространства.
12.5 ХЕМОРЕЦЕПЦИЯВзаимоотношения рыб с внешней средой объединяют в две группы факторов: абиотические и биотические. Физические и химические свойства воды, действующие на рыб, называют абиотическими факторами.
Восприятие животными химические вещества с помощью рецепторов — одна из форм реакции организмов на воздействие внешней среды. У водных животных специализированные рецепторы контактируют с веществами, находящимися в растворенном состоянии, поэтому характерное для наземных животных четкое разделение на рецепторы обоняния, воспринимающие летучие вещества, и рецепторы вкуса, воспринимающие вещества, находящиеся в твердом и жидком состоянии, у водных животных не проявляется. Однако морфологически и функционально органы обоняния у рыб достаточно хорошо обособлены. Основываясь на отсутствии специфичности в функционировании, локализации и связи с нервными центрами, принято объединять вкус и общее химическое чувство понятием «химический анализатор», или «необонятельная хеморецепция».
ОРГАН ОБОНЯНИЯ относится к группе химических рецепторов. Органы обоняния рыб расположены в ноздрях, расположеных впереди каждого глаза, форма и величина которых меняется в зависимости от экологии. Представляют собой простые ямки со слизистой оболочкой, пронизанные разветвлением нервов, ведущие в слепой мешок с чувствительными клетками идущих от обонятельной доли головного мозга.
У большинства рыб каждая из ноздрей поделена перегородкой на автономные передние и задние носовые отверстия. В некоторых случаях, носовые отверстия одинарные. В онтогенезе носовые отверстия у всех рыб первоначально одинарны, т.е. не поделены перегородкой на переднюю и заднюю ноздри, которые обособляются лишь на более поздних стадиях развития.
Расположение ноздрей у различных видов рыб зависит от их образа жизни и развития других органов чувств. У рыб с хорошо развитым зрением носовые отверстия расположены на верхней стороне головы между глазом и концом рыла. У селяхше ноздри располагаются на нижней стороне и приближены к ротовому отверстию.
Относительная величина ноздрей тесно связана со скоростью движения рыбы. У рыб, плавающих медленно, ноздри сравнительно больше, причем перегородка между передним и задним носовыми отверстиями имеет вид вертикально расположенного щитка, направляющего воду обонятельную капсулу. У рыб быстрых, носовые отверстия чрезвычайно маленькие, так как при больших скоростях встречного обтекающего конька, вода в носовой капсуле смываются достаточно быстро, через относительно маленькие отверстия передних ноздрей. У придонных рыб, у которых роль обоняния в общей системе рецепции очень существенна, передние носовые отверстия вытягиваются в виде трубочек и приближаются к ротовой щели или даже свешиваются с верхней челюсти в низ, это имеет место, у Typhleotris, Anguilla, Mnreana и др.
Пахучие вещества, растворенные в воде, попадают на слизистую оболочку обонятельной области, раздражают окончания обонятельных нервов, отсюда сигналы поступают в головной мозг.
 Посредством обоняния рыбы получают информацию об изменениях внешней среды, различают пищу, находят свою стаю, партнеров во время нереста, обнаруживают хищников, вычисляют добычу. На коже у некоторых видов рыб расположены клетки, которые при ранении кожи выделяют в воду «вещество страха», которое является для других рыб сигналом об опасности. Рыбы активно используют химическую информацию для подачи сигналов тревоги, предупреждения об опасности, привлечения особей противоположного пола. Особенно важен этот орган для рыб, обитающих в мутной воде, где наряду с осязательной и звуковой информацией рыбы активно используют и обонятельную систему. Обоняние оказывает большое влияние и на работу многих органов и систем организма, тонизируя или угнетая их. Известны группы веществ, положительно (аттраканты) или отрицательно (репелленты) действующие на рыб. Обоняние тесно связано с другими органами чувств: вкуса, зрения и равновесия.
Посредством обоняния рыбы получают информацию об изменениях внешней среды, различают пищу, находят свою стаю, партнеров во время нереста, обнаруживают хищников, вычисляют добычу. На коже у некоторых видов рыб расположены клетки, которые при ранении кожи выделяют в воду «вещество страха», которое является для других рыб сигналом об опасности. Рыбы активно используют химическую информацию для подачи сигналов тревоги, предупреждения об опасности, привлечения особей противоположного пола. Особенно важен этот орган для рыб, обитающих в мутной воде, где наряду с осязательной и звуковой информацией рыбы активно используют и обонятельную систему. Обоняние оказывает большое влияние и на работу многих органов и систем организма, тонизируя или угнетая их. Известны группы веществ, положительно (аттраканты) или отрицательно (репелленты) действующие на рыб. Обоняние тесно связано с другими органами чувств: вкуса, зрения и равновесия.
В различные времена года обонятельные ощущения у рыб не одинаковы, они обостряются весной и летом, особенно в теплую погоду.
 Рыбы, ведущие ночной образ жизни (угорь, налим, сом) обладают высокоразвитым обонянием. Обонятельные клетки этих рыб способны реагировать сотые доли концентраций аттракантов и репеллентов.
Рыбы, ведущие ночной образ жизни (угорь, налим, сом) обладают высокоразвитым обонянием. Обонятельные клетки этих рыб способны реагировать сотые доли концентраций аттракантов и репеллентов.
 Рыбы способны почувствовать разведение в пропорции один к миллиарду экстракта из мотыля, карась чувствует аналогичную концентрацию нитробензола, более высокие концентрации менее привлекательны для рыб. Для обонятельного эпителия стимуляторами служат аминокислоты, некоторые из них или же их смеси имеют сигнальное значение для рыб. Например, угорь находит моллюска по выделяемому им комплексу, состоящему из 7 аминокислот. Позвоночные животные ориентируются на смесь основных запахов: мускусный, камфорный, мятный, эфирный, цветочный, острый и гнилой.
Рыбы способны почувствовать разведение в пропорции один к миллиарду экстракта из мотыля, карась чувствует аналогичную концентрацию нитробензола, более высокие концентрации менее привлекательны для рыб. Для обонятельного эпителия стимуляторами служат аминокислоты, некоторые из них или же их смеси имеют сигнальное значение для рыб. Например, угорь находит моллюска по выделяемому им комплексу, состоящему из 7 аминокислот. Позвоночные животные ориентируются на смесь основных запахов: мускусный, камфорный, мятный, эфирный, цветочный, острый и гнилой.
Рецепторы обоняния у рыб, как и у прочих позвоночных, парные и расположены на передней части головы. Лишь у крутлоротых непарные. Обонятельные рецепторы находятся на слепом углублении — ноздре, дно которой выстлано обонятельным эпителием, располагающимся на поверхности складок. Складки, расходясь радиально от центра, образуют обонятельную розетку.
У разных рыб обонятельные клетки расположены на складках по-разному: сплошным слоем, разреженно, на гребнях или в углублении. Ток воды, несущий молекулы пахучих веществ, попадает к рецептору через переднее отверстие, часто отделеное от выходного заднего отверстия лишь складкой кожи. Однако у некоторых рыб входное и выходное отверстия заметно разделены и далеко стоят друг от друга. Передние (входные) отверстия у ряда рыб (угорь, налим) находятся близко к концу рыла и снабжены кожными трубочками. Считается, что этот признак свидетельствует о значительной роли обоняния в поисках пищевых объектов. Движение воды в обонятельной ямке может создаваться либо движением ресничек на поверхности выстилки, либо сокращением и расслаблением стенок специальных полостей — ампул, либо в результате движения самой рыбы.
Рецепторные обонятельные клетки, имеющие биполярную форму, относятся к категории первичных рецепторов, т. е. сами регенерируют импульсы, содержащие информацию о раздражителе и передают их по отросткам к нервным центрам. Периферический отросток обонятельных клеток направляется к поверхности рецепторного слоя и заканчивается расширением — булавой, на апикальном конце которой имеется пучок волосков или микроворсинок. Волоски пронизывают слой слизи на поверхности эпителия и способны к движениям.
Обонятельные клетки окружены опорными клетками, которые содержат овальные ядра и многочисленные гранулы разного размера. Здесь же расположены базальные клетки, не содержащие секреторных гранул. Центральные отростки рецепторных клеток, не имеющие миелиновой оболочки, пройдя базальную мембрану эпителия, образуют пучки до нескольких сотен волокон, окруженные мезаксоном шванновской клетки, причем тело одной клетки может охватывать много пучков. Пучки сливаются в стволики, образующие обонятельный нерв, соединяющийся с обонятельной луковицей.
Строение обонятеельной выстилки у всех позвоночных сходно (рис. 95), что свидетельствует о сходстве механизма контактной рецепции. Однако сам этот механизм еше не вполне ясен. Одна из них связывает возможности распознавания запахов, т. е. молекул пахучих веществ, с избирательной специфичностью индивидуальных рецепторов к запахам. Такова стереохимическая гипотеза Эймура. согласно которой, имеется семь типов активных мест на обонятельных клетках, а молекулы веществ обладающих сходными запахами, имеют одинаковую форму активных частей, которые подходят к активным точкам рецептора, как » ключ» к замку. Другие гипотезы связывают возможность различения запахов с различиями в распределении веществ, адсорбируемых слизью обонятельной выстилки, по ее поверхности. Ряд исследователей полагают, что распознавание запахов обеспечивают эти два механизма, дополняя друг друга.
Ведущая роль — в обонятельной рецепции принадлежат, волоскам и булаве обонятельной клетки, которые обеспечивают специфическое взаимодействие молекул одорантов с мембраной клетки и перевод эффекта взаимодействия в форму электрического потенциала. Как уже было сказано, аксоны обонятельных рецепторных клеток образуют обонятельный нерв, входящий в обонятельную луковицу, являющуюся первичным центром обонятельного рецептора.
Обонятельная луковица относится, как считает А. А. Заварзин, к экранным структурам. Для нее характерно расположение элементов в виде последовательных слоев, причем нервные элементы связаны между собой не только в пределах слоя, но и между слоями. Таких слоев выделяют обычно три: слой обонятельных клубочков с межклубочковыми клетками, слой вторичных нейронов с митральными и кисточковыми клетками, гранулярный слой.
Информацию в высшие обонятельные центры у рыб передают вторичные нейроны и клетки гранулярного слоя. Наружная часть обонятельной луковицы состоит из волокон обонятельного нерва, контакт которых с дендритами вторичных нейронов происходит в обонятельных клубочках, где наблюдается ветвление тех и других окончаний. В одном обонятельном клубочке сходятся несколько сотен волокон обонятельного нерва. Слои обонятельной луковицы обычно расположены концентрически, но у некоторых видов рыб (щука), они лежат последовательно в рострокаудальном направлении.
Обонятельные луковицы рыб анатомически хорошо обособлены и бывают двух типов: сидячие, прилегающие к переднему мозгу; стебельчатые, расположенные сразу за рецепторами (очень короткие обонятельные нервы).
У тресковых, обонятельные луковицы связаны с передним мозгом длинными обонятельными трактами, которые представлены медиальным и латеральным пучками, оканчивающимися в ядрах переднего мозга.
Обоняние как способ получения информации об окружающем мире имеет для рыб весьма существенное значение. По степени развития обоняния рыб, как и других животных, обычно делят на макросматиков и микросматиков. Это деление связанно с различной широтой спектра воспринимаемых запахов.
У макресматиков органы обоняния способны воспринимать большое количество различных запахов, т. е. обоняние используется ими в более разнообразных ситуациях.
Микросматики воспринимают обычно неболышое количество запахов — в основном особей своего вида и половых партнерш. Типичным представителем макросматиков являётся обыкновенний угорь, микросматиков — щука, трехиглая колюшка. Для восприятия запаха иногда, видимо, достаточно попадания на обонятельный рецептор, нескольких молекул вещества.
Обоняние может играть руководящую роль в поисках пищи, особенно у ночных и сумеречных хищников, таких, как угорь. С помощью обоняния рыбы могут воспринимать партнеров по стае, находить особей другого пола в период размножения. Например, гольян может различать среди особей своего вида партнера. Рыбы одного вида способны воспринимать химические соединения, выделяемые кожей других рыб при ранении.
Изучение миграций проходных лососей показало, что на этапе входа в нерестовые реки они отыскивают именно ту реку, где сами вывелись из икры, ориентируясь по запаху воды, запечатленному в памяти на этапе молоди (рис.96). Источниками запаха, видимо, являются виды рыб, постоянно обитающие в реке. Эта способность была использована для направления мигрирующих производителей в определенный участок. Молодь кижуча выдерживалась в растворе морфолина с концентрацией 0~5 М, а затем, после их возвращения в период нереста в родную реку, привлекалась тем же раствором к определенному месту в водоеме.

Рис. 96. Биотоки обонятельного мозга лосося при орошении обонятельных ямок; 1, 2 — дистиллированной водой; 3 — водой из родной реки; 4, 5, 6 — водой из чужих озер.
Рыба обладает обонянием, которое сильнее развито у нехищных рыб. Щука, например, при поисках пищи не пользуется обонянием. При ее быстром броске за добычей обоняние не может играть существенной роли. Другой хищник — окунь при передвижениях в поисках пищи обычно плавает тихо, подбирая со дна всевозможные личинки, он обонянием в данном случае пользуется как органом, наводящим на пищу.
Орган вкуса имеется почти у всех рыб, вкусовые ощущения передаются большинству из них через губы и рот. Поэтому рыба не всегда проглатывает схваченный корм, особенно если он не пришелся ей по вкусу.
Вкус — это ощущение, возникающее при действии на орган вкуса пищевых и некоторых не пищевых веществ. Орган вкуса тесно связан с органом обоняния и относится к группе химических рецепторов. Вкусовые ощущения у рыб появляются при раздражении чувствительных, осязательных клеток — вкусовых сосочек или так называемых вкусовых почек, луковиц, расположенных в ротовой полости в виде микроскопических вкусовых клеток, на усиках, по всей поверхности тела, особенно на кожных выростах. (Рис.97)
Главными восприятиями вкусовых ощущении являются четыре составляющих: кислое, сладкое, соленое и горькое. Остальные виды вкуса представляют собой комбинации этих четырех ощущений, причем вкусовые ощущения у рыб могут вызывать только вещества, растворенные в воде.
Минимально ощутимая разница в концентрации растворов веществ порог различия — постепенно ухудшается при переходе от слабых к более сильным концентрациям. К примеру, однопроцентный раствор сахара обладает практически максимально сладким вкусом, и дальнейшее увеличение его концентрации не изменяет вкусового ощущения.
Появление вкусовых ощущений может быть вызвано действием на рецептор неадекватных раздражителей, например, постоянного электрического тока. При длительном соприкосновении какого-либо вещества с органом вкуса постепенно притупляется его восприятие, в конце концов, это вещество покажется рыбе совершенно безвкусным происходит адаптация.
Вкусовой анализатор также может влиять на некоторые реакции организма, на деятельность внутренних органов. Установлено, что рыбы реагируют практически на все имеющие вкус вещества и обладают при этом поразительно тонким вкусом. Положительные или отрицательные реакции рыб определяются их образом жизни и, прежде всего, характером их питания. Положительные реакции на сахар свойственны животным, питающимся растительной и смешанной пищей. Ощущение горечи у большинства живых существ вызывает отрицательную реакцию, но не у тех, которые питаются насекомыми.
Рис.97 . Расположение вкусовых почек на теле сома показано точками. Каждая точка означает 100 вкусовых почек
Механизм вкусовой рецепции.Четыре основных вкусовых ощущения — сладкое, горькое, кислое и соленое — воспринимаются благодаря взаимодействию молекул вкусовых веществ с четырьмя молекулами белков. Комбинации этих типов и создают конкретные вкусовые ощущения. У большинства рыб вкус играет роль контактной рецепции, поскольку пороги вкусовой чувствительности сравнительно высоки. Но у некоторых рыб вкус может приобретать функции дистантного рецептора. Так, пресноводный сомик с помощью вкусовых рецепторов способен локализовать пищу на расстоянии около 30 длин тела. При отключении вкусовых рецепторов эта способность исчезает. С помощью общей химической чувствительность рыбы способны улавливать изменения солености до 0,3% концентрации отдельных солей, изменения концентрации растворов органических кислот (лимонная) до 0,0025 М (0,3 г/л), изменения рН порядка 0,05—0,07 концентрации углекислоты до 0,6 г/л.
Необонятельная хеморецепция у рыб осуществляется вкусовыми почками и свободными окончаниями блуждающего, тройничного и некоторых спинномозговых нервов. Структура вкусовых почек сходна у всех классов позвоночных. У рыб они обычно имеют овальную форму и состоят из 30—50 вытянутых клеток, апикальные концы которых образуют канал. К основанию клеток подходят окончания нерва. Это типичные вторичные рецепторы. Они располагаются в ротовой полости, на губах, жабрах, в глотке, на коже головы и туловища, на усиках и плавниках. Их количество варьирует от 50 до сотен тысяч и зависит, как и их расположение, в большей мере от экологии, нежели от вида. Размеры, количество и распределение вкусовых почек характеризует степень развития вкусовой рецепции конкретного вида рыб. Вкусовые почки передней части рта и кожи инервируются волокнами возвратной ветви лицевого нерва, а слизистой рта и жабр — волокнами языкоглоточиого и блуждающего нервов. В иннервации вкусовых рецепторов, участвуют также тройничный и смешанные нервы.
§
Химические соединения воспринимаются также нервными окончаниями — сенсорными аппаратами в виде цилиндров, каналов, веретен, кустиков. Установлено, что передняя часть, боковой линии и ампулы Лоренцйни хрящевых рыб являются рецепторами, воспринимающими изменения солености воды.
Центры регуляции вкусовой и общей химической чувствительности находятся в продолговатом мозге, в вагусной доле или в обособленной доле лицевого нерва (у карповых и силурид). Эти центры имеют связи с ядрами таламуса и гипоталамуса, а также среднего мозга.
Велика сигнальная роль различных органических соединений, выделяемых рыбами в окружающую среду. Некоторые из них являются специфическими раздражителями, определяющими многие формы поведения рыб: пищевое, межвидовое, стайное, репродуктивное. К таким веществам относятся аминокислоты, амины — прежде всего алифатические, пептиды, органические кислоты и др. Часто их состав весьма сложен, а действуют они в столь малых концентрациях, что возникают трудности с определением их химической природы. Они могут восприниматься как обонятельной системой, так и вкусовыми и общехимическими рецепторами. Расшифровка состава этих натуральных химических раздражителей, обеспечивающих рыбам детальную и многообразную информацию об окружающей среде, может дать в руки исследователей и практиков эффективное средство управления поведением рыб в самых разнообразных ситуациях.
Чувствительность рыб к различным веществам неодинакова. Некоторые вещества рыбы могут различать в концентрациях менее 0,1 мкг/л, а другие они совсем не воспринимают
12.6 ЭЛЕКТРОРЕЦЕПЦИЯ Рыбы обитают в среде, хорошо проводящей электричество, поэтому электрорецепция играет в их жизни важную роль. Чувствительностью к электричеству обладают в той или иной мере всерыбы, однако она различна в зависимости от развития рецепторов. Рыбы, не имеющие специальных чувствительных образований, способны ощущать ток свободными нервными окончаниями, а также по воздействию на другие органы чувств, в частности на органы боковой линии. Такие рыбы ощущают импульсы тока с напряжением в десятые и сотые доли вольта на 1 см: карп — 0,06—0,1 В/см; форель — 0,02— 0,08В/см; карась — 0,008—0,015 В/см.
Рыбы, имеющие электрорецепторы, обладают высокой чувствительностью к электрическим полям — от десятых долей милливольта до сотых долей микровольта: минога — 0,1 мкВ/см; химера — 0,2 мкВ/см; морские акулы и скаты-до 0,01 мкВ/см.
Электрорецепторы (рис.98.) берут начало от органов боковой линии, располагаются на голове, спине, брюшной части тела, но отсутствуют в области расположения электрических органов. У акул и скатов роль электрорецепторов играют ампулы Лоренцини, у мормирид -бугорчатые органы, у сомов — ямкообразные органы. Органы Лоренцини представляют собой длинные, иногда по нескольку сантиметров, трубки, открывающиеся наружу порами. В слепом конце трубки имеется расширение — ампула, в которой находятся чувствительные клетки, погруженные в желеобразную массу. Бугорчатые органы имеют вид возвышений на коже, они состоят из чувствительных и опорных клеток.
Ампулярные органы работают, как гальванометры, реагируя на изменение силы тока, а бугорчатые — как осциллографы, воспринимая изменение напряжения внешнего электрического поля.
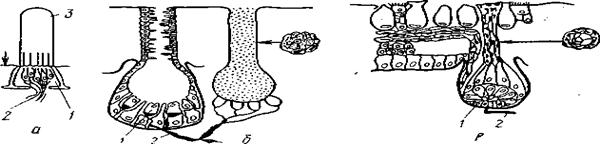
Рис. 98. Кожные электрорецепторы рыб:
а-ординарные рецепторы боковой линии; б — ампулярные рецепторы; в — бугорковые органы; 1 — рецепторные клетки; 2 — нерв; 3 — купула
Рыбы с чувствительными рецепторами (акулы, мормириды) способны реагировать на электробиологические явления, например биотоки дыхательных мышц кормовых организмов, закопавшихся в грунт. Кроме того, существует реальная возможность использования магнитно-гидродинамических эффектов, возникающих при движении рыб в магнитном поле земли. Ощутимые для чувствительных рецепторов электрические токи возникают при скорости плавания рыб около 1 м/с.1
При действии на рыб постоянного тока отмечаются следующие реакции: ориентировочная реакция — вздрагивание при включении и выключении (0,03—0,3 В/см); оборонительная реакция — беспокойство, попытки уйти из зоны действия тока при увеличении напряжения до 0,1—1,5 В/см; анодная реакция — рыба поворачивается вдоль силовых линий и плывет в сторону анода (0,3—2,5 В/см); потеря равновесия, подвижности, электронаркоз при дальнейшем повышении напряжения. В морской воде электрический ток действует сильнее, чем в пресной. Сильные токи убивают рыбу.
В поле переменного тока наблюдаются примерно те же две первые фазы действия, что и в поле постоянного тока, при дальнейшем повышении напряженности появляется осцилло – таксис, когда рыба располагается поперек силовых линий и развивается электронаркоз.
Последствия действия переменного тока длятся дольше, чем последствия действия постоянного тока. Действие импульсного тока зависит от амплитуды, частоты, формы и продолжительности импульсов. Действие регистрируется видимой реакцией как вздрагивания, но при анализе кардиограммы или при выработке условных рефлексов удается обнаружить на порядок более низкие пороги, чем при определении видимой реакции. Видимая реакция мальков карпа наблюдается при 2 В.
При воздействии ядовитых веществ как фенол возбудимость повышается и порог понижается до 1 В, а при воздействии солевых растворов возбудимость понижается и порог возрастает до 3,5— 4 В. Оптимальная длительность импульсов для стимулирования реакции вздрагивания находится около 5 мс, максимальная сила мышечной реакции достигается в случае контактного приложения электродов, при достижении силы тока 4—6 В.
При частом электрическом стимулировании отмечается прывыкание организма рыб, так приблизительно после 100 стимулов сила реакции уменьшается вдвое в результате привыкания и электроутомления. После 300—600 импульсов рыба перестает реагировать на электрическую стимуляцию и становится неподвижной. Такое электроутомление наступает быстрее у больных и истощенных рыб, что можно использовать для оценки общего состояния организма рыбы.
Импульсный ток затрудняет плавание. При частотах, в 1,5—2 раза превышающих максимальные частоты плавательных движений (50 Гц), наступает тетанус мышц. Судорожные напряжения мышц при высокой частоте стимуляции продолжаются до тех пор, пока не произойдет электроутомление нервно-мышечной системы и рыба не станет неподвижной (300— 600 импульсов, длительность 5—10 сек. при частоте 50 Гц). Такая неподвижность рыб используется при ловле рыб электросачками «Баклан» и электротралами. В этих устройствах используются электроимпульсы высокой частоты.
12.7 ТЕРМОРЕЦЕПЦИЯ Терморецепторы, по-видимому, рассеяны по всей поверхности тела рыбы. Реагировать на изменения температуры могут многие чувствительные элементы, в том числе ампулы Лоренцини. Рыбы обладают хорошо развитой способностью определять разность температур. Пороги чувствительности разных видов рыб колеблются между десятыми и сотыми долями градуса. Наиболее чуткими к изменениям температуры являются некоторые донные морские рыбы. Например, морские собачки, подкаменщики, бельдюги, мерланги и бычки реагируют на повышение температуры воды на 0,03°С. Разницу в 0,05°С, улавливают некоторые морские и пресноводные рыбы — сайда, дорш, пинацр, камбала, зеленушка, карась, колюшка. Радужная форель и ушастый окунь могут реагировать на различие температуры в 0,1°С. гольян и сомик реагируют только на повышение или понижение температуры на 1°С.
Отношение рыб к температуре характеризуется их термопреферендумом – предпочитаемой нормальной температурой, которая, естественно, зависит от привычной температуры. Конечный термопреферендум трески 9°С, радужной форели 16-20°С, тропической тиляпии 28-29°С. Термопреферендум некоторых карповых рыб различен зимой и летом. Лещ, синец, плотва и другие и обитатели наших рек летом стремятся к температуре 27°С, а зимой — к температуре 2-3°С, при которой лучше проходит зимовка.
При минусовых температурах воздуха над открытой поверхностью водоемов частички воды, граничащие с атмосферой, охлаждаются до точки замерзания и, если при этом имеется течение или дует ветер, происходит перемещение охлажденной воды с подлежащими слоями. Охлажденные частицы воды, соприкасаясь с подводными предметами, выпадают на них в виде кристаллов льда. Эти предметы являются зонами кристаллизации. Если в этот период рыба попадает в соприкосновение с охлажденными частицами воды, она сама становится такой зоной и уже не в состоянии покинуть эту зону, впадая в состояние оцепенения, и гибнет. В периоды, когда температура приближается к нулевой отметке и переходит ее, но еще остаются открытые водные пространства, рыба испытывает сильный страх. В это время она должна прятаться в глубокие ямы и места, где полностью отсутствует течение, что и наблюдается в действительности.
Условия для образования внутриводного льда могут иметь место не глубокой осенью, они могут складываться и весной, и в зимний период, особенно в водоемах с течением.
Под действием ветра лед вибрирует, в результате чего прилегающие к нижней кромке льда переохлажденные частички воды могут отрываться и, попадая в нижние слои воды, охлаждать и постепенно истончать теплый слой воды, такое явление может способствовать возникновению зимних заморов рыб, как говорят, вызванных кислородным голоданием. Но известно, что чем холоднее вода, тем больше в ней растворяется кислорода. Процессы гниения растений затормаживаются, а толщина льда не может иметь сильно отрицательного значения. Заморы происходят в суровые зимы, когда создаются условия не только для истончения, но и полного исчезновения теплого слоя воды. Образование внутриводного льда возможно также и подольдом, если с нижней поверхности охлажденного льда происходит как бы «слизывание»переохлажденных частиц воды течением. Эти частицы также могут найти себе точки кристаллизации.
Видимо, образование внутриводного льда может происходить при низких сезонных температурах воды и при водозаборах воды у плотин, а в периоды оттепелей, отсутствуют условия образования внутриводного льда и дальнейшего охлаждения слоя воды. Описанное явление следует рассматривать как составляющую более широкой проблемы в жизни рыб, связанной с сезонным охлаждением воды. Для многих видов рыб охлажденной водой в наших широтах зимой следует считать воду с температурой ниже 4° С.
Охладившись до этой температуры, вода опускается на дно, заполняя постепенно водоем от дна до поверхности. При дальнейшем понижении температуры над теплым слоем образуются слои воды, охлажденной до 3, 2, 1°С, и поверхностный слой с температурой 0°С, в котором происходит наращивание льда, перемешивания слоев воды подольдом не происходит. Физиологические функции организма рыб при переходе к зимним условиям существования резко снижаются, а при определенных температурах убывают до какого-то нижнего предела.
При встрече с температурой воды ниже какого-то предела (у каждого вида рыбы и даже возрастной группы одного вида эти температурные пределы различны) рыбы впадают в коматозное состояние. Многие рыбы, например, сом, карп, карась, линь и другие, впадают в спячку при температурах ниже 4 °С. А такие рыбы как форель, хариус и другие, активны и при очень охлажденной воде. Налим же после становления льда устремляется к самой холодной воде, к слою, граничащему со льдом. Видимо, температура, близкая к О °С, необходима ему для подготовки к нересту, а нереститься он начинает при температуре 0,5 °С.
Рыбы обладают экологической терморегуляцией, т.е.всегда стремятся в сторону оптимальных для них температур, Это прежде всего места, где температура равна 4 °С, а также места с более высокой температурой, что возможно вследствие подогрева воды за счет других источников — ключей, ручьев, теплоэлектростанций и т. п.
Температура в значительной степени оказывает влияние и на кормовую базу рыб. Из всего комплекса факторов, влияющих на поведение рыб, температурный в условиях наших северных широт являются доминирующим, определяющим их поведение. В широтах, прилегающих к экватору, температурный фактор является второстепенным, там главенствует гравитационный фактор. Конечно же, другие факторы, и прежде всего световой, имеют очень важное значение в жизни рыб, как и в жизни всех живых существ. Световой фактор руководит суточными биоритмами рыб и в какой-то мере совместно с другими факторами влияет на их сезонное поведение. Основой для проявления как температурного, так и светового факторов является Солнце — единственный источник тепловой и световой энергии для Земли. В условиях зимы, т.е. при очень холодной погоде, как и в летнюю жару, при температуре выше 30° С, о влиянии температуры на рыб следует помнить всегда.
Рыбы — хладнокровные существа, температура тела которых может повышаться только за счет активизации ее движения, но не более, чем на 1-2 градуса. Интервал температур, в котором могут существовать рыбы, очень ограничен в плюсовом диапазоне, от температуры несколько выше нуля до 20-30° С. Для каждого вида рыб имеется определенный температурный интервал, характеризуемый минимумом, оптимумом и максимумом температуры.
Температурные пределы для одного и того же вида рыб несколько меняются в зависимости от того, в каком цикле развития в данный момент находятся рыбы, а также от их пола и предшествующих температурных условий. Каждый биологический процесс на различных стадиях развития рыб характеризуется своим температурным оптимумом. Например, весенний посленерестовый жор или жор рыб по перволедью протекают при температурах, далеких от оптимальных. Природа в первом случае позаботилась о том, чтобы рыбы восполнили потерю сил, а во втором случае активизировали подготовку к зиме.
Рыбы еще используют так называемую экологическую терморегуляцию, они перемещаются в сторону наиболее благоприятных для них в данный период температур. Когда температура воды в местах обитания рыб опускается или поднимается за пределы переносимого интервала, рыбы покидают эти места (это так называемая сезонная миграция), либо впадают в зимнюю или летнюю спячку, если они лишены возможности мигрировать. Рыбы обладают очень высокой чувствительностью к смене температуры и чувствуют ее изменение даже в пределах, измеряемых тысячными долями градуса.
Температура воды зависит от температуры воздуха и изменяется медленнее вследствие большой теплоемкости воды. Суточные изменения температуры воды незначительны, хотя они и повторяют общий ход изменения температуры воздуха, отставая от последней на 4° С.Известно, что в разные годы в водоемы могут поступать большие или меньшие объемы холодной воды, нагрев которых, естественно, требует большего или меньшего времени. Даже при холодной, но солнечной погоде, например, дно водоема прогревается лучше, особенно на мелководье, и, следовательно, отдает воде больше тепла.
Предел температур, способствующих интенсивному питанию большинства видов рыб колеблется примерно от 10°до25°С. В жару большинство рыб (окунь, лещ, плотва, язь, карп и другие) прячутся в тени, голавль поднимается на поверхность, налим чувствует себя нормально при температурах до 12ْ°С, уже при 15° С он впадает в летнюю спячку.В этом плане показательны наблюдения исследователей за поведением молоди белого амура: наибольшая ее активность происходит при 20-22°С, при 12° С активность снижается вдвое, а при 10° С рыбы прекращают реагировать на внешние раздражители. У карпа наибольшая прибавка в весе замечена при 20-28°С, при 14° С резко снижается потребление пищи, а при 1-2° С он впадает в малоподвижное состояние. При 20°С. суточный ритм жизни леща близок к максимальному.
Важность температурного фактора в наших условиях заключена не только в том, что он является определяющим в поведении рыб, но и в том, что он лежит в основе развития растительного и животного мира как животной и растительной кормовой базы рыб. Растения развиваются в зависимости от температурного фактора и ориентируются на суммы активных (положительных) температур. Корма животного и растительного происхождения для рыб не менее чувствительны, чем сами рыбы к изменению температуры. Рыбы, как и любые другие живые существа, испытывают воздействие температурного фактора, имеющего, с одной стороны, строго сезонный характер, а с другой — зависящего от других факторов, к которым следует отнести и различные погодные явления.
Температура напрямую влияет и на кислородный режим воды, с ее повышением содержание кислорода в воде уменьшается.
Если действие какого-либо конкретного фактора, явления или суммарного действия нескольких факторов ведет к изменению температуры воды в сторону оптимального для жизни рыб значения, то рыбы активизируются.
Тактильная чувствительность.Помимо выше указанных органов восприятия, у некоторых рыб на плавниковых лучах имеются специальные органы ощупывания в виде различных усиков и мясистых выростов. У некоторых, например у гурами, одна пара плавников видоизменилась и превратилась в чрезвычайно подвижные щупальцы, которыми рыба ощупывает различные предметы и животных, включая и себе подобных.

Рис.99 Типы усов у рыб
Приспособления, обеспечивающие работу тактильных и вкусовых рецепторов, во внешнем строении рыбы проявляются в возникновении различно рода придатков, на которых эти рецепторы располагаются. Сюда относятся удлиненные специализированные лучи плавников – брюшных (Trichogastor, Ptrophyllum, Ophidion, Cadidae),грудных(Triglidae), спинного (Chanliadus) и др. а также усики расположенные на голове. (Рис. 99)
Большей частью указанные придатки несут на себе как тактильные, так и вкусовые рецепторы. С морфологической стороны рассматриваемые образования весьма разнообразны. Лучи плавников могут быть удлиненные чрезвычайно подвижны, могут быть поправлены в любую сторону. Удлинение и обособлении лучей плавников обусловлено не только тем, что на них располагаются тактильные и вкусовые рецепторы. Но и тем, что они отчасти являются органом, при помощи которого рыбы передвигается по субстрату, примером, могут служить обособленные лучи грудных плавников Trigla. Усики также весьма неодинаковы у разных видов, как по числу, так и по форме и размерам. Усики, удлиненные лучи плавников. Свойственные преимущественно придонным и придонно- пелагическими рыбам, у которых значение этих органов чувств в общей системе рецепции наиболее велико. Кроме того, существует много специальных органов — луковиц, дисков, телец, спиралей, воспринимающих и передающих в нервные центры сигналы о механических воздействиях на ткань. У рыб большое количество тактильных рецепторов находится в коже и плавниках.
Механорецепторы находятся у рыб в сухожилиях, что позволяет рыбе регистрировать направление струй воды, касание предметов, анализировать свою позу. Характерных для высших позвоночных чувствительных веретен, расположенных в мышцах, у рыб нет.
При низком подпороговом воздействии электрический потенциал чувствительной клетки несколько повышается, но настоящие импульсные разряды появляются при превышении пороговой степени воздействия.
Таблица 22. Упругие колебания и их восприятие рыбами
| Явление | Частоты, гц | Воспринимающие структуры |
| Вибрация | 0,1—16 | Тактильные рецепторы кожи |
| Звук низкой частоты | 16—100 | Боковая линия, плавательный пузырь |
| Звук низкой частоты | 100—1000 | Внутреннее ухо (лабиринт) |
| Звук высокой частотой | 1000—15000 | Внутреннее ухо при наличии Веберово аппарата веберова аппарата |
| Ультразвук | Выше 15000 | Не ощущаются при низкой интенсивности. |
Некоторые механорецепторы дают только один разряд в начале приложения раздражителя, другие дают пульсирующие разряды в течение всего периода механической деформации. Усиление воздействия, например, давления, растяжения, учащают эти разряды. Упругие периодические колебания среды — звуки и вибрации — воспринимаются разными органами рыб (табл.22).
12.8 ЗВУКИ, ИЗДАВАЕМЫЕ РЫБАМИ.Рыбы издают много разнообразных звуков. Звуки представляют собой колебания среды и являются результатами тех или иных движений тела рыбы или его частей. Некоторые звуки издаются рыбами специально для общения друг с другом. Есть звуки, сопровождающие жизнедеятельность рыб,— это упругие волны и колебания, возникающие при плавании, питании, захвате воздуха. выдавливании воздуха из плавательного пузыря.
Спектр звуков, связанных с движением, охватывает область частот от 20 Гц до 5 кГц. Эти звуки носят шумовой характер и отличаются невысоким давлением (1005—205 Па) на расстоянии 1 м от рыб. Низкочастотные явления связаны с плавательными колебаниями тела, частота которых, как известно, обычно находится в пределах 4—20 Гц. Высокочастотные звуки связаны с турбулентными возмущениями среды.
Некоторые звуки связаны с газообменом. Многие виды рыб (карповые, лососевые, лабиринтовые) пользуются звуками для дыхания и наполнения газового пузыря атмосферным воздухом. Проталкивание воздуха через узкие отверстия (воздушный канал пузыря, пищевод, кишечник, анус) вызывает звуки, напоминающие слабый писк.

 При захвате добычи хищными рыбами (судаком, щукой и др.) раздается звук, напоминающий хлопок, удар или звук откупориваемой бутылки. Эти звуки гидродинамические, низкочастотные, с максимальной амплитудой в области 100 Гц,. Мирные и всеядные рыбы при питании издают звуки, напоминающие причмокивание или приглушенное цоканье, что может вызываться шевелением гравия, отрыванием от субстрата растительности, хватанием моллюсков и ракообразных. При этом возникают звуки с максимумом в области 400-600 Гц и силой до 30 дБ. В результате перетирания пищи зубами также возникают хрусты и скрежеты, зависящие от качества корма. Довольно громкие звуки типа «бум-бум-бум» возникают при разжевывании карпом сухого зерна. При этом явно возникает резонанс в плавательном пузыре. Скрежеты, щелчки и хрусты, или стридуляции, возникают при трении челюстных и глоточных зубов, а также сочленений жестких плавниковых лучей. Некоторые из этих звуков произвольные и имеют коммуникативное значение. Таковы звуки, издаваемые при шевелении передними плавниками касатки-скрипуна и некоторых спинорогов.
При захвате добычи хищными рыбами (судаком, щукой и др.) раздается звук, напоминающий хлопок, удар или звук откупориваемой бутылки. Эти звуки гидродинамические, низкочастотные, с максимальной амплитудой в области 100 Гц,. Мирные и всеядные рыбы при питании издают звуки, напоминающие причмокивание или приглушенное цоканье, что может вызываться шевелением гравия, отрыванием от субстрата растительности, хватанием моллюсков и ракообразных. При этом возникают звуки с максимумом в области 400-600 Гц и силой до 30 дБ. В результате перетирания пищи зубами также возникают хрусты и скрежеты, зависящие от качества корма. Довольно громкие звуки типа «бум-бум-бум» возникают при разжевывании карпом сухого зерна. При этом явно возникает резонанс в плавательном пузыре. Скрежеты, щелчки и хрусты, или стридуляции, возникают при трении челюстных и глоточных зубов, а также сочленений жестких плавниковых лучей. Некоторые из этих звуков произвольные и имеют коммуникативное значение. Таковы звуки, издаваемые при шевелении передними плавниками касатки-скрипуна и некоторых спинорогов.
Контрольные вопросы.
1.Что такое анализаторы их функциональное назначение?
2.Строение зрительного анализатора, мехенизм восприятия света и цвета.
3.Охарактеризуйте структуру и функции сетчатки
4.Какие фотохимические реакции и электрические явления протекают в сетчатке?
5. Какова острота зрения у рыб?
6.Строение слухового анализатора, мехенизм восприятия звука.
7.Строение вкусового анализатора, мехенизм восприятия вкуса.
8. Каково строение обонятельного анализатора, мехенизм восприятия запахов у костистых и хрящевых рыб?
9.Острота и чувствительность обонянии у рыб
10.Класификация и теории запахов.
11.Болевая, температурная, барометрическая и тактильная рецепция и их значение в жизнедеятельности рыб.
12.Обьясните механизм возникновения возбуждения рецептрного аппарата органов восприятия.
13 Механизм издавания звуков рыбами.
14.Обьясните механизм нервной и гуморальной регуляции работ органов и систем восприятии.
Таблица 23 — Вещества, привлекающие рыб к пище и наживке
|
Таблица.24- Величина концентрации некоторых веществ, воспринимаемая рыбами
| Вещество | Концентрация | Рыба | Примечания | |||||
| Сахароза Сахароза | 0,24 г/л 7,5 мг/л | Форель Пескарь | В 18 раз чувствительнее человека В100 раз | |||||
| Хлористый натрий | 2 мг/л | То же | Различает | |||||
| Ион меди | 0,1 мкг/’л | лосось | Избегает | |||||
| Ион меди | 100 мкг/л | Черный окунь | Повышенная частота дыхания | |||||
| Ион меди | 10 мкг/л | Карась | Различает | |||||
| Ион меди | 50 мкг/л Разведение | То же Лосось | Привлекает с То же | |||||
| Сок планктона | Разведение 1 : 25 000 | То же | » | |||||
| Эвгенол (пахучее | | Карп | Избегает | |||||
| Ион цинка | 5 — 7 мкг/л | Форель | То же | |||||
| Хлор молекулярный | 1 мкг/л | То же | Слабо избегает | |||||
| Хлор молекулярный | 10 мкг/л | » | Заметно избегает | |||||
| Хлор молекулярный | 1000 мкг/л | » | Сильно избегает | |||||
СЛОВАРЬ ТЕРМИНОВ — глоссарии