

Рекомендации
- ^Пратт CA, Voet D, Voet JG (2002). Основы модернизации биохимии. Нью-Йорк: Вили. С. 264–266. ISBN 0-471-41759-9.
- ^Фридман, Мортон (2008). Принципы и модели биологического транспорта. Springer. ISBN 978-0387-79239-2.
- ^ абcКленин, Константин В .; Мерлиц, Хольгер; Ланговски, Йорг; Ву, Чен-Сюй (2006). «Облегченная диффузия ДНК-связывающих белков». Письма с физическими проверками. 96 (1): 018104. arXiv:физика / 0507056. Дои:10.1103 / PhysRevLett.96.018104. ISSN 0031-9007. PMID 16486524.
- ^ абБауэр М., Мецлер Р. (2022). «Модель облегченной диффузии in vivo». PLoS ONE. 8 (1): e53956. arXiv:1301.5502. Bibcode:2022PLoSO … 853956B. Дои:10.1371 / journal.pone.0053956. ЧВК 3548819. PMID 23349772.
- ^ абHammar, P .; Leroy, P .; Махмутович, А .; Marklund, E. G .; Berg, O.G .; Эльф, Дж. (2022). «Lac Repressor показывает облегченную диффузию в живых клетках». Наука. 336 (6088): 1595–1598. Bibcode:2022Научный … 336.1595H. Дои:10.1126 / science.1221648. ISSN 0036-8075. PMID 22723426.
- ^Brackley CA, Cates ME, Marenduzzo D (сентябрь 2022 г.). «Внутриклеточное облегченное распространение: искатели, крауддеры и блокаторы». Phys. Rev. Lett. 111 (10): 108101. arXiv:1309.1010. Bibcode:2022PhRvL.111j8101B. Дои:10.1103 / PhysRevLett.111.108101. PMID 25166711.
- ^ абБенишу О., Шевалье С., Мейер Б., Войтурие Р. (январь 2022 г.). «Облегченная диффузия белков на хроматин». Phys. Rev. Lett. 106 (3): 038102. arXiv:1006.4758. Bibcode:2022PhRvL.106c8102B. Дои:10.1103 / PhysRevLett.106.038102. PMID 21405302.
- ^Крейцер, Ф. (1970). «Облегченная диффузия кислорода и его возможное значение; обзор». Физиология дыхания. 9 (1): 1–30. Дои:10.1016/0034-5687(70)90002-2. ISSN 0034-5687. PMID 4910215.
- ^Jacquez JA, Kutchai H, Daniels E (июнь 1972 г.). «Диффузия кислорода, облегчаемая гемоглобином: межфазные эффекты и эффекты толщины»(PDF). Респир физиол. 15 (2): 166–81. Дои:10.1016/0034-5687(72)90096-5. HDL:2027.42/34087. PMID 5042165.
- ^Рубинов С.И., Дембо М. (апрель 1977 г.). «Облегченная диффузия кислорода гемоглобином и миоглобином». Биофиз. J. 18 (1): 29–42. Bibcode:1977BpJ …. 18 … 29R. Дои:10.1016 / S0006-3495 (77) 85594-X. ЧВК 1473276. PMID 856316.
- ^Kreuzer F, Hoofd LJ (май 1972 г.). «Факторы, влияющие на облегчение диффузии кислорода в присутствии гемоглобина и миоглобина». Респир физиол. 15 (1): 104–24. Дои:10.1016/0034-5687(72)90008-4. PMID 5079218.
- ^ абВиттенберг JB (январь 1966 г.). «Молекулярный механизм диффузии кислорода, облегченной гемоглобином». J. Biol. Chem. 241 (1): 104–14. PMID 5901041.
- ^Мюррей Дж. Д., Вайман Дж. (Октябрь 1971 г.). «Облегченная диффузия. Случай окиси углерода». J. Biol. Chem. 246 (19): 5903–6. PMID 5116656.
- ^Торенс Б (1993). «Облегченные переносчики глюкозы в эпителиальных клетках». Анну. Преподобный Physiol. 55: 591–608. Дои:10.1146 / annurev.ph.55.030193.003111. PMID 8466187.
- ^ абКаррутерс, А. (1990). «Облегченная диффузия глюкозы». Физиологические обзоры. 70 (4): 1135–1176. Дои:10.1152 / Physrev.1990.70.4.1135. ISSN 0031-9333. PMID 2217557.
Способы транспорта веществ через мембрану.
Большинство процессов жизнедеятельности, таких, как всасывание, выделение, проведение нервного импульса, мышечное сокращение, синтез АТФ, поддержание постоянства ионного состава и содержания воды связано с переносом веществ через мембраны. Этот процесс в биологических системах получил название транспорта. Обмен веществ между клеткой и окружающей её средой происходит постоянно. Механизмы транспорта веществ в клетку и из неё зависят от размеров транспортируемых частиц. Малые молекулы и ионы транспортируются клеткой непосредственно через мембрану в форме пассивного и активного транспорта.
Пассивный транспорт осуществляется без затрат энергии, по градиенту концентрации путем простой диффузии, фильтрации, осмоса или облегченной диффузии.
Диффузия – проникновение веществ через мембрану по градиенту концентрации (из области, где их концентрация выше, в область, где их концентрация ниже); этот процесс происходит без затрат энергии вследствие хаотического движения молекул. Диффузный транспорт веществ (вода, ионы) осуществляется при участии интегральных белков мембраны, в которых имеются молекулярные поры (каналы, через которые проходят растворенные молекулы и ионы), либо при участии липидной фазы (для жирорастворимых веществ). С помощью диффузии в клетку проникают растворенные молекулы кислорода и углекислого газа, а также яды и лекарственные препараты.
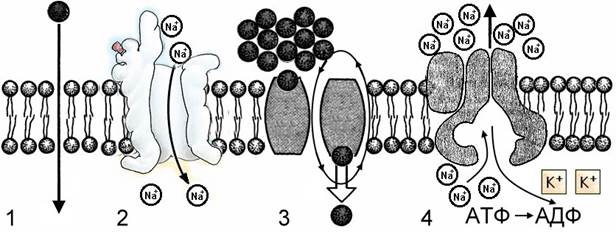
Рис. Виды транспорта через мембрану.1 – простая диффузия; 2 – диффузия через мембранные каналы; 3 – облегченная диффузия с помощью белков-переносчиков; 4 – активный транспорт.
Облегченная диффузия. Транспорт веществ через липидный бислой с помощью простой диффузии совершается с малой скоростью, особенно в случае заряженных частиц, и почти не контролируется. Поэтому в процессе эволюции для некоторых веществ появились специфические мембранные каналы и мембранные переносчики, которые способствуют повышению скорости переноса и, кроме того, осуществляют селективный транспорт. Пассивный транспорт веществ с помощью переносчиков называется облегченной диффузией. Специальные белки-переносчики (пермеаза) встроены в мембрану. Пермеазы избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану. При этом частицы перемещаются быстрее, чем при обычной диффузии.
Осмос – поступление в клетки воды из гипотонического раствора.
Фильтрация — просачивание веществ поры в сторону меньших значений давления. Примером фильтрации в организме является перенос воды через стенки кровеносных сосудов, выдавливание плазмы крови в почечные канальцы.
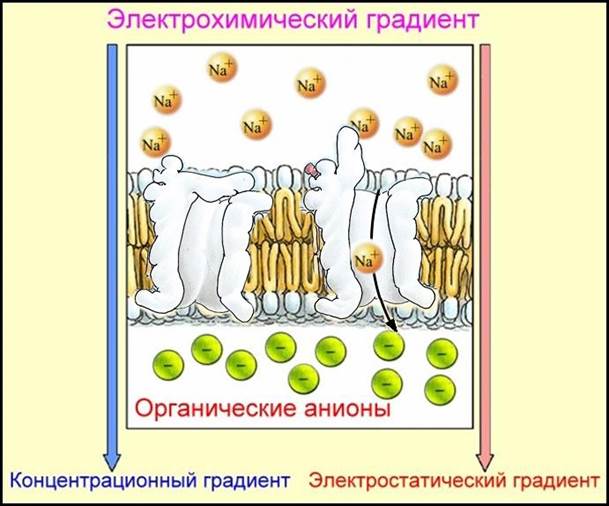
Рис. Движение катионов по электрохимическому градиенту.
Активный транспорт. Если бы в клетках существовал только пассивный транспорт, то концентрации, давления и др. величины вне и внутри клетки сравнялись бы. Поэтому существует другой механизм, работающий в направлении против электрохимического градиента и происходящий с затратой энергии клеткой. Перенос молекул и ионов против электрохимического градиента, осуществляемый клеткой за счет энергии метаболических процессов, называется активным транспортом.Он присущ только биологическим мембранам. Активный перенос вещества через мембрану происходит за счет свободной энергии, высвобождающейся в ходе химических реакций внутри клетки. Активный транспорт в организме создает градиенты концентраций, электректрических потенциалов, давлений, т.е. поддерживает жизнь в организме.
Активный транспорт заключается в перемещении веществ против градиента концентрации с помощью транспортных белков (порины, АТФ-азы и др.), образующих мембранные насосы, с затратой энергии АТФ (калий-натриевый насос, регуляция концентрации в клетках ионов кальция и магния, поступление моносахаридов, нуклеотидов, аминокислот). Изучены 3 основные системы активного транспорта, которые обеспечивают перенос ионов Na, K, Ca, H через мембрану.
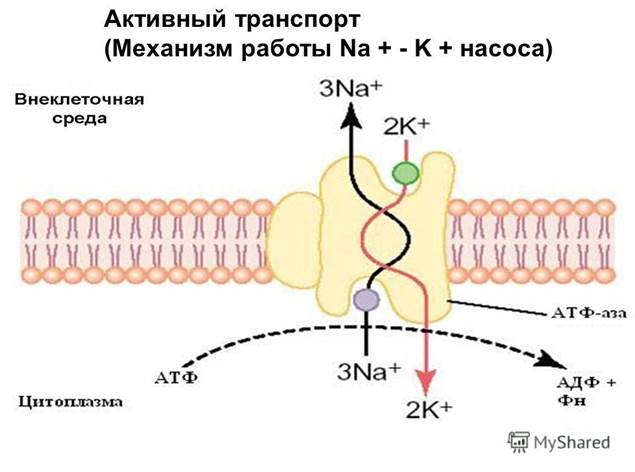
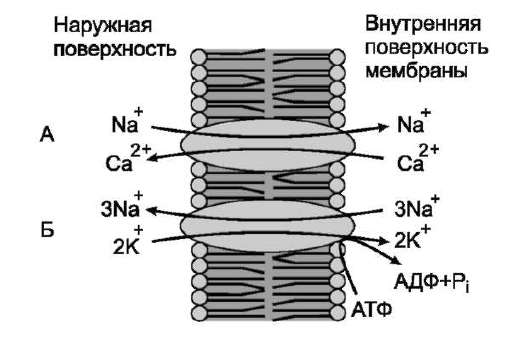
Механизм. Ионы К и Na неравномерно распределены по разные стороны мембраны: концентрация Na снаружи > ионов K , а внутри клетки K > Na . Эти ионы диффундируют через мембрану по направлению электрохимического градиента, что приводит к его выравниванию. Na-K насосы входят в состав цитоплазматических мембран и работают за счет энергии гидролиза молекул АТФ с образованием молекул АДФ и неорганического фосфата Фн: АТФ=АДФ Фн.Насос работает обратимо: градиенты концентраций ионов способствуют синтезу молекул АТФ из мол-л АДФ и Фн: АДФ Фн=АТФ.
Na /К -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как «K », так и «Na ». За один цикл работы насос выводит из клетки три «Na » и заводит два «К » за счет энергии молекулы АТФ. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки.
Через мембрану могут переноситься не только отдельные молекулы, но и твердые тела (фагоцитоз ), растворы ( пиноцитоз ). Фагоцитоз –захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и пиноцитоз – захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Образующиеся пиноцитозные вакуоли имеют размеры от 0,01 до 1-2 мкм. Затем вакуоль погружается в цитоплазму и отшнуровывается. При этом стенка пиноцитозной вакуоли полностью сохраняет структуру породившей ее плазматической мембраны.

Если вещество транспортируется внутрь клетки, то такой вид транспорта называется эндоцитозом (перенос в клетку путем прямого пино-или фагоцитоза), если наружу, то – экзоцитозом (перенос из клетки путем обратного пино — или фагоцитоза). В первом случае на наружной стороне мембраны образуется впячивание, которое постепенно превращается в пузырек. Пузырек отрывается от мембраны внутри клетки. Такой пузырек содержит в себе транспортируемое вещество, окруженное билипидной оболочкой (везикулой). В дальнейшем везикула сливается с какой-нибудь клеточной органеллой и выпускает в неё своё содержимое. В случае экзоцитоза процесс происходит в обратной последовательности: везикула подходит к мембране с внутренней стороны клетки, сливается с ней и выбрасывает своё содержимое в межклеточное пространство.
Пиноцитоз и фагоцитоз – принципиально сходные процессы, в которых можно выделить четыре фазы: поступление веществ путем пино-или фагоцитоза, их расщепление под действием ферментов выделяемых лизосомами, перенос продуктов расщепления в цитоплазму (вследствие изменения проницаемости мембран вакуолей) и выделение наружу продуктов обмена. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
§
Цитоплазма – обязательная составная часть клетки. В ней происходят сложные и разнообразные процессы синтеза, дыхания, роста, ей присущи явления раздражимости и наследственности, т.е. все те свойства, которые характеризуют жизнь.
Цитоплазма представляет собой вязкую прозрачную бесцветную массу с удельным весом 1,04 – 1,06. Свет преломляет чуть сильнее воды. Цитоплазма упруга, эластична, с водой не смешивается. Во многих клетках можно наблюдать ее движение: в клетках с одной крупной центральной вакуолью – круговое (циклоз), в клетках со многими вакуолями и тяжами цитоплазмы между ними – струйчатое. Ток цитоплазмы вовлекает в движение клеточные органоиды.
Цитоплазма дифференцирована на бесструктурную массу – гиалоплазму и оформленные образования – клеточные органоиды. Гиалоплазма(цитоплазматический матрикс) – сложная коллоидная система, образованная белками, нуклеиновыми кислотами, углеводами, водой и другими веществами. В зависимости от физиологического состояния и воздействия внешней среды гиалоплазма может находиться в виде золя (жидкости) или геля (более упругого плотного вещества). Гиалоплазма является внутренней средой клетки, где протекают реакции внутриклеточного обмена.
В гиалоплазме клеток, между ядерной оболочкой и цитоплазматической мембраной, расположен цитоскелет – механический каркас цитоплазмы. Он представляет собой сложную трехмерную сеть филаментов (белковых трубочек): микрофиламентов (6 – 8 нм), образованных белком актином; промежуточных волокон (10 нм), состоящих из разных фибриллярных белков (цитокератинов и др.); микротрубочек (около 25 нм), построенных из тубулина и способных сокращаться. Цитоскелет выполняет различные функции:
— определяет форму клетки;
— упорядочивает размещение всех структурных компонентов клетки;
— участвует в различных движениях самой клетки (при делении) и во внутриклеточном перемещении органоидов и отдельных соединений;
— микротрубочки непосредственно участвуют в образовании веретена деления и растягивании хромосом при митозе и мейозе;
— по микротрубочкам органоиды передвигаются в те места клетки, где они нужны в данный момент;
— микрофиламенты взаимодействуют с микротрубочками и обеспечивают амёбоидное движение одноклеточных организмов;
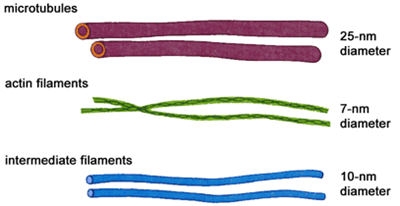
Микротрубочки содержатся во всех эукариотических клетках и представляют собой полые неразветвленные цилиндры, диаметр которых не превышает 30 нм, а толщина стенки — 5 нм. В длину они могут достигать нескольких микрометров. Легко распадаются и собираются вновь. Стенка микротрубочек в основном построена из спирально уложенных субъединиц белка тубулина. Считают, что роль матрицы (организатора микротрубочек) могут играть центриоли, базальные тельца жгутиков и ресничек, центромеры хромосом. Функции микротрубочек: выполняют опорную функцию; придают клетке определенную форму; образуют веретено деления; обеспечивают расхождение хромосом к полюсам клетки; отвечают за перемещение клеточных органелл; принимают участие во внутриклеточном транспорте, секреции, формировании клеточной стенки; являются структурным компонентом ресничек, жгутиков, базальных телец и центриолей.
Микрофиламенты представлены нитями диаметром 6 нм, состоящими из белка актина, близкого к актину мышц. Актин составляет 10-15% общего количества белка клетки. В большинстве животных клеток образуется густая сеть из актиновых филаментов и связанных с ними белков под самой плазматической мембраной. Эта сеть придает поверхностному слою клетки механическую прочность и позволяет клетке изменять свою форму и двигаться. Помимо актина, в клетке обнаруживаются и нити миозина. Однако количество их значительно меньше. Благодаря взаимодействию актина и миозина происходит сокращение мышц.
Микрофиламенты связаны с движением всей клетки либо ее отдельных структур внутри нее. В некоторых случаях движение обеспечивается только актиновыми филаментами, в других — актином вместе с миозином.
Микротрабекулярная система представляет собой сеть из тонких фибрилл — трабекул (перекладин), в точках пересечения или соединения концов которых располагаются рибосомы. Микротрабекулярная система — динамичная структура: при изменении условий она может распадаться и вновь собираться. Функции микротрабекулярной решетки: служит опорой для клеточных органелл; осуществляет связь между отдельными частями клетки; направляет внутриклеточный транспорт.
Элементы цитоскелета очень динамичны: в определенных участках клетки при изменении внешних и внутренних условий могут распадаться на отдельные молекулы и вновь собираться. Образующиеся молекулы белка переходят в раствор в составе гиалоплазмы, изменяя агрегатное состояние гиалоплазмы с гелеобразного (гель) на жидкое (золь). При полимеризации белковых субъединиц и сборке микротрубочек и микрофиламентов наблюдается обратный процесс.
Функции гиалоплазмы:
1) является внутренней средой клетки, в которой происходят многие химические процессы;
2) объединяет все клеточные структуры и обеспечивает химическое взаимодействие между ними;
3) определяет местоположение органоидов в клетке;
4) обеспечивает внутриклеточный транспорт веществ (аминокислот, сахаров и др.) и перемещение органоидов (движение хлоропластов в растительных клетках);
5) является зоной перемещения молекул АТФ;
6) определяет форму клетки.
Цитоплазма – сложная химическая многокомпонентная система, содержащая 75-86% воды, 10-20% белков, 2-3% липидов, 1-2% углеводов, 1% минеральных солей. Это суммарный и приблизительный состав цитоплазмы, который не отражает сложности ее химической структуры.
В цитоплазме в растворенном состоянии содержится большое количество свободных аминокислот и нуклеотидов, множество промежуточных продуктов, возникающих при синтезе и распаде молекул. Также обнаруживается большое количество ионов Na , K , Mg2 , Cl—, HCO32-, HPO42- и др.
§
Органоиды – постоянные и обязательные компоненты клеток; специализированные участки цитоплазмы клетки, имеющие определенную структуру и выполняющие определенные функции в клетке. Различают органоиды общего и специального назначения.
Органоиды общего назначения имеются в большинстве клеток (эндоплазматическая сеть, митохондрии, пластиды, комплекс Гольджи, лизосомы, вакуоли, клеточный центр, рибосомы). Органоиды специального назначения характерны только для специализированных клеток (миофибриллы, жгутики, реснички, сократительные и пищеварительные вакуоли). Органоиды (за исключением рибосом и клеточного центра) имеют мембранное строение.
Эндоплазматическая ретикулум(ЭПР) – это разветвленная система соединенных между собой полостей, трубочек и каналов, образованных элементарными мембранами и пронизывающая всю толщу клетки. Открыта в 1943 г. Портером. Особенно много каналов эндоплазматической сети в клетках с интенсивным обменом веществ. В среднем объем ЭПС составляет от 30% до 50% общего объема клетки. ЭПС лабильна. Форма внутренних лакун и кана
лов, их размер, расположение в клетке и количество изменяются в процессе жизнедеятельности. Развита сильнее в животных клетка. ЭПС морфологически и функционально связана с пограничным слоем цитоплазмы, ядерной оболочкой, рибосомами, комплексом Гольджи, вакуолями, образуя вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и передвижения веществ внутри клетки. Вблизи эндоплазматической сети накапливаются митохондрии и пластиды.
Выделяют две разновидности ЭПС: шероховатую и гладкую. На мембранах гладкой (агранулярной) ЭПС локализованы ферменты систем жирового и углеводного синтеза: здесь происходит синтез углеводов и почти всех клеточных липидов. Мембраны гладкой разновидности эндоплазматической сети преобладают в клетках сальных желез, печени (синтез гликогена), в клетках с большим содержанием питательных веществ (семена растений). На мембране шероховатой (гранулярной) ЭПС располагаются рибосомы, где осуществляется биосинтез белков. Часть синтезируемых ими белков включается в состав мембраны эндоплазматической сети, остальные поступают в просвет ее каналов, где преобразуются и транспортируются в комплекс Гольджи. Особенно много шероховатых мембран в клетках желез и нервных клетках.
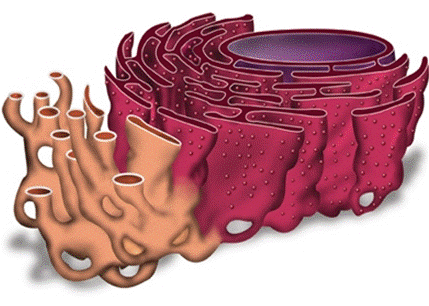
Рис. Шероховатый и гладкий эндоплазматический ретикулум.
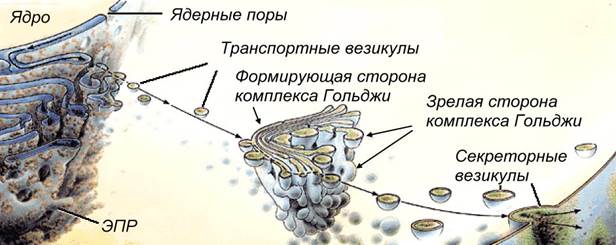
Рис. Транспорт веществ по системе ядро – эндоплазматический ретикулум (ЭПР) – комплекс Гольджи.
Функции эндоплазматической сети:
1) синтез белков (шероховатая ЭПС), углеводов и липидов (гладкая ЭПС);
2) транспорт веществ, как поступивших в клетку, так и вновь синтезируемых;
3) деление цитоплазмы на отсеки (компартменты), что обеспечивает пространственное разделение ферментных систем, необходимое для их последовательного вступления в биохимические реакции.
Митохондрии – присутствуют практически во всех типах клеток одно- и многоклеточных организмов (за исключением эритроцитов млекопитающих). Число их в разных клетках варьирует и зависит от уровня функциональной активности клетки. В клетке печени крысы их около 2500, а в мужской половой клетке некоторых моллюсков – 20 – 22. Их больше в грудной мышце летающих птиц, чем в грудной мышце нелетающих.
Митохондрии имеют форму сферических, овальных и цилиндрических телец. Размеры составляют 0,2 – 1,0 мкм и диаметре и до 5 — 7 мкм в длину.
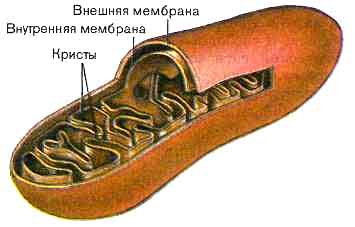
Рис. Митохондрия.
Длина нитевидных форм достигает 15-20 мкм. Снаружи митохондрии ограничены гладкой наружной мембраной, сходной по составу с плазмалеммой. Внутренняя мембрана образует многочисленные выросты – кристы – и содержит многочисленные ферменты, АТФ-сомы (грибовидные тела), участвующие в процессах трансформации энергии питательных веществ в энергию АТФ. Количество крист зависит от функции клетки. В митохондриях мышц крист очень много, они занимают всю внутреннюю полость органоида. В митохондриях эмбриональных клеток кристы единичны. В растительных выросты внутренней мембраны чаще имеют форму трубочек. Полость митохондрии заполнена матриксом, в котором содержатся вода, минеральные соли, белки-ферменты, аминокислоты. Митохондрии имеют автономную белоксинтезирующую систему: кольцевую молекулу ДНК, различные виды РНК и более мелкие, чем в цитоплазме рибосомы.
Митохондрии тесно связаны мембранами эндоплазматической сети, каналы которой часто открываются прямо в митохондрии. При повышении нагрузки на орган и усилении синтетических процессов, требующих затраты энергии, контакты между ЭПС и митохондриями становятся особенно многочисленными. Число митохондрий может быстро увеличиваться путем деления. Способность митохондрий к размножению обусловлена присутствием в них молекулы ДНК, напоминающей кольцевую хромосому бактерий.
Функции митохондрий:
1) синтез универсального источника энергии – АТФ;
2) синтез стероидных гормонов;
3) биосинтез специфических белков.
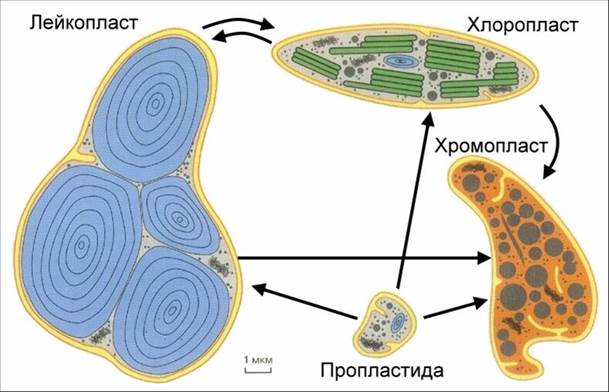
Пластиды – органоиды мембранного строения, характерные только для растительных клеток. В них происходят процессы синтеза углеводов, белков и жиров. По содержанию пигментов их делят на три группы: хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют относительно постоянную эллиптическую или линзовидную форму. Размер по наибольшему диаметру составляет 4 – 10 мкм. Количество в клетке колеблется от нескольких единиц до нескольких десятков. Их размер, интенсивность окраски, количество и расположение в клетке зависят от условий освещения, вида и физиологического состояния растений.
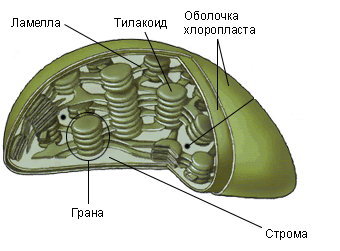
Рис. Хлоропласт, строение.
Это белково-липоидные тела, состоящие на 35-55% из белка, 20-30% — липидов, 9% — хлорофилла, 4-5% каратиноидов, 2-4% нуклеиновых кислот. Количество углеводов варьирует; обнаружено некоторое количество минеральных веществ Хлорофилл – сложный эфир органической двухосновной кислоты – хлорофиллина и органических спиртов – метилового (СН3ОН) и фитола (С20Н39ОН). У высших растений в хлоропластах постоянно присутствуют хлорофилл а – имеет сине-зеленую окраску, и хлорофилл b – желто-зеленую; причем содержание хлорофилла, а в несколько раз больше.
Кроме хлорофилла в состав хлоропластов входят пигменты — каротин С40Н56 и ксантофилл С40Н56О2 и некоторые другие пигменты (каратиноиды). В зеленом листе желтые спутники хлорофилла маскируются более яркой зеленой окраской. Однако осенью, при листопаде, у большинства растений хлорофилл разрушается и тогда обнаруживается присутствие в листе каратиноида – лист становится желтым.
Хлоропласт одет двойной оболочкой, состоящей из наружной и внутренней мембран. Внутреннее содержимое – строма – имеет ламеллярное (пластинчатое) строение. В бесцветной строме выделяют граны – окрашенные в зеленые цвет тельца, 0,3 – 1,7 мкм. Они представляют собой совокупность тилакоидов – замкнутых телец в виде плоских пузырьков или дисков мембранного происхождения. Хлорофилл в виде мономолекулярного слоя располагается между белковым и липидным слоями в тесной связи с ними. Пространственное расположение молекул пигментов в мембранных структурах хлоропластов является весьма целесообразным и создает оптимальные условия для наиболее эффективного поглощения, передачи и использования лучистой энергии. Липиды образуют безводные диэлектрические слои мембран хлоропласта, необходимые для функционирования электронно-транспортной цепи. Роль звеньев цепи переноса электронов выполняют белки (цитохромы, пластохиноны, ферредоксин, пластоцианин) и отдельные химические элементы – железо, марганец и др. Количество гран в хлоропласте от 20 до 200. Между гранами, связывая их друг с другом, располагаются ламеллы стромы. Ламеллы гран и ламеллы стромы имеют мембранное строение.
Внутренне строение хлоропласта делает возможным пространственное разобщение многочисленных и разнообразных реакций, составляющих в своей совокупности содержание фотосинтеза.
Хлоропласты, как и митохондрии, содержат специфическую РНК и ДНК, а также более мелкие рибосомы и весь молекулярный арсенал, необходимый для биосинтеза белка. У этих органоидов имеется достаточное для обеспечения максимальной активности белоксинтезирующей системы количество и-РНК. Вместе с тем в них содержится и достаточно ДНК для кодирования определенных белков. Они размножаются делением, путем простой перетяжки.
Установлено, что хлоропласты могут изменять свою форму, размеры и положение в клетке, т. е. способны самостоятельно двигаться (таксис хлоропластов). В них обнаружено два типа сократительных белков, за счет которых, очевидно, и осуществляется активное движение этих органоидов в цитоплазме.
Хромопласты широко распространены в генеративных органах растений. Они окрашивают лепестки цветков (лютика, георгина, подсолнечника), плоды (томатов, рябины, шиповника) в желтый, оранжевый, красный цвета. В вегетативных органах хромопласты встречаются значительно реже.
Окраска хромопластов обусловлена присутствием каратиноидов – каротина, ксантофилла и ликопина, которые в пластидах находятся в различном состоянии: в виде кристаллов, липоидного раствора или в соединении с белками.
Хромопласты, по сравнению с хлоропластами, имеют более простое строение – в них отсутствует ламеллярная структура. Химический состав также отличен: пигменты – 20–50%, липиды до 50%, белки – около 20%, РНК – 2-3%. Это свидетельствует о меньшей физиологической активности хлоропластов.
Лейкопласты не содержат пигментов, они бесцветны. Эти самые мелкие пластиды имеют округлую, яйцевидную или палочковидную форму. В клетке они часто группируются вокруг ядра.
Внутренне структура, еще менее дифференцирована по сравнению с хлоропластами. В них осуществляется синтез крахмала, жиров, белков. В соответствии с этим выделяют три вида лейкопластов – амилопласты (крахмал), олеопласты (растительные масла) и протеопласты (белки).
Возникают лейкопласты из пропластид, с которыми они сходны по форме и строению, а отличаются лишь размерами.
Все пластиды генетически связаны друг с другом. Они образуются из пропластид – мельчайших бесцветных цитоплазматических образований, сходных по внешнему виду с митохондриями. Пропластиды находятся в спорах, яйцеклетках, в эмбриональных клетках точек роста. Непосредственно из пропластид образуются хлоропласты (на свету) и лейкопласты (в темноте), а из них развиваются хромопласты, являющиеся конечным продуктом в эволюции пластид в клетке.
Комплекс Гольджи – впервые был обнаружен в 1898 г. Итальянским ученым Гольджи в животных клетках. Это система внутренних полостей, цистерн (5-20), располагающихся сближено и параллельно друг другу, и крупных и мелких вакуолей. Все эти образования имеют мембранное строение и являются специализированными участками эндоплазматической сети. В животных клетках комплекс Гольджи развит лучше, чем в растительных; в последних он называется диктиосомы.
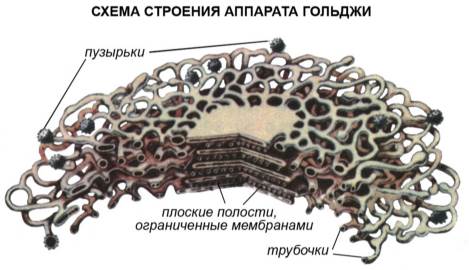
Рис. Строение комплекса Гольджи.
Попадающие в пластинчатый комплекс белки и липиды, подвергаются различным преобразованиям, накапливаются, сортируются, упаковываются в секреторные пузырьки и транспортируются по назначению: к различным структурам внутри клетки или за пределы клетки. Мембраны комплекса Гольджи также синтезируют полисахариды и образуют лизосомы. В клетках молочных желез комплекс Гольджи участвует в образовании молока, а в клетках печени – желчи.
Функции комплекса Гольджи:
1) концентрация, обезвоживание и уплотнение синтезированных в клетке белков, жиров, полисахаридов и веществ, поступивших извне;
2) сборка сложных комплексов органических веществ и подготовка их к выведению из клетки (целлюлоза и гемицеллюлоза у растений, гликопротеины и гликолипиды у животных);
3) синтез полисахаридов;
4) образование первичных лизосом.
Лизосомы — небольшие овальные тельца диаметром 0,2-2,0 мкм. Центральное положение занимает вакуоль, содержащая 40 (по разным данным 30-60) гидролитических ферментов, способных в кислой среде (рН 4,5-5) расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и другие вещества.
Вокруг этой полости располагается строма, одетая снаружи элементарной мембраной. Расщепление веществ при помощи ферментов называется лизисом, поэтому органоид назван лизосомой. Образование лизосом происходит в комплексе Гольджи. Первичные лизосомы приближаются непосредственно к пиноцитозным или фагоцитозным вакуолям (эндосомам) и изливают свое содержимое в их полость, образуя вторичные лизосомы (фагосомы), внутри которых и происходит переваривание веществ. Продукты лизиса через мембрану лизосом поступают в цитоплазму и включаются в дальнейший обмен веществ. Вторичные лизосомы с остатками непереваренных веществ называются остаточными тельцами. Примером вторичных лизосом являются пищеварительные вакуоли простейших.
Функции лизосом:
1) внутриклеточное переваривание макромолекул пищи и чужеродных компонентов, поступающих в клетку при пино- и фагоцитозе, обеспечивая клетку дополнительным сырьем для биохимических и энергетических процессов;
2) при голодании лизосомы переваривают некоторые органоиды и на какое-то время пополняют запас питательных веществ;
3) разрушение временных органов эмбрионов и личинок (хвост и жабры у лягушки) в процессе постэмбрионального развития;
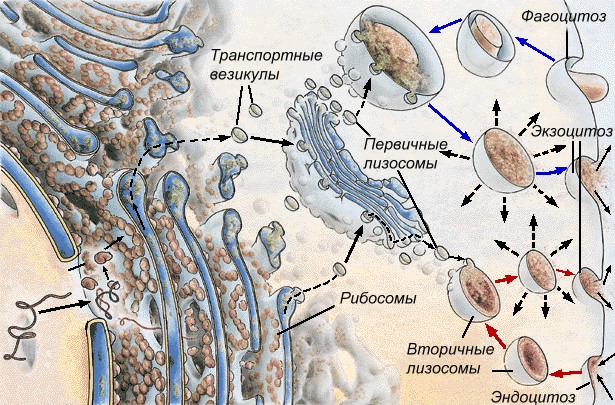
Рис. Образование лизосом
Вакуоли – полости в цитоплазме растительных клеток и протист, заполненные жидкостью. Имеют форму пузырьков, тонких канальцев и другую. Вакуоли образуются из расширений эндоплазматической сети и пузырьков комплекса Гольджи как тончайшие полости, затем по мере роста клетки и накопления продуктов обмена объем их увеличивается, а количество сокращается. Развитая сформировавшаяся клетка имеет обычно одну большую вакуоль, занимающую центральное положение.
Вакуоли растительных клеток заполнены клеточным соком, который представляет собой водный раствор органических (яблочная, щавелевая, лимонная кислоты, сахара, инулин, аминокислоты, белки, дубильные вещества, алкалоиды, глюкозиды) и минеральных (нитраты, хлориды, фосфаты) веществ.
У протист встречаются пищеварительные вакуоли и сократительные.
Функции вакуолей:
1) хранилища запасных питательных веществ и вместилища выделений ( у растений);
2) определяют и поддерживают осмотическое давление в клетках;
3) обеспечивают внутриклеточное пищеварение у протист.
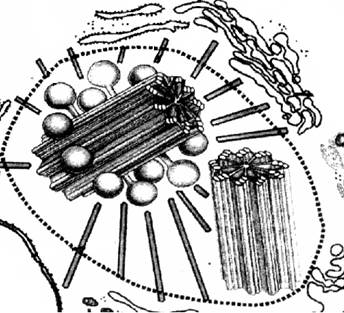
Рис. Клеточный центр.
Клеточный центр обычно находится вблизи ядра и состоит из двух центриолей, расположенных перпендикулярно друг другу и окруженных лучистой сферой. Каждая центриоль представляет собой полое цилиндрическое тельце длиной 0,3-0,5 мкм и длиной 0,15 мкм, стенка которого образована 9 триплетами микротрубочек. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем.
Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу. Они формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками, являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам. В клетках высших растений клеточный центр центриолей не имеет.
Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли.
Центросома характерна для клеток животных, некоторых грибов, водорослей, мхов и папоротников.
Функции клеточного центра:
1) образование полюсов деления и формирование микротрубочек веретена деления.
Рибосомы — мелкие сферические органоиды, от 15 до 35 нм. Состоят из двух субъединиц большой (60S) и малой (40S). Содержат около 60% белка и 40% рибосомальной РНК. Молекулы рРНК образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка. Субъединицы рибосом образуются в ядрышках. и через поры в ядерной оболочке поступают в цитоплазму, где располагаются либо на мембране ЭПА, либо на наружной стороне ядерной оболочки, либо свободно в цитоплазме. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка. По сравнению с митохондриями, пластидами, клетками прокариот рибосомы в цитоплазме эукариотических клеток крупнее. Могут объединяться по 5-70 единиц в полисомы.
Функции рибосом:
1) участие в биосинтезе белка.
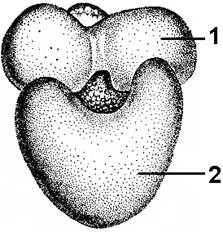
Рис. 287. Рибосома: 1 — малая субъединица; 2 — большая субъединица.
Реснички, жгутики – выросты цитоплазмы, покрытые элементарной мембраной, под которой находится 20 микротрубочек, образующих 9 пар по периферии и две одиночные в центре. У основания ресничек и жгутиков расположены базальные тельца. Длина жгутиков достигает 100 мкм. Реснички – это короткие – 10-20 мкм – жгутики. Движение жгутиков винтовое, а ресничек – веслообразное. Благодаря ресничкам и жгутикам передвигаются бактерии, протисты, ресничные, перемещаются частицы или жидкости (реснички мерцательного эпителия дыхательных путей, яйцеводов), половые клетки (сперматозоиды).
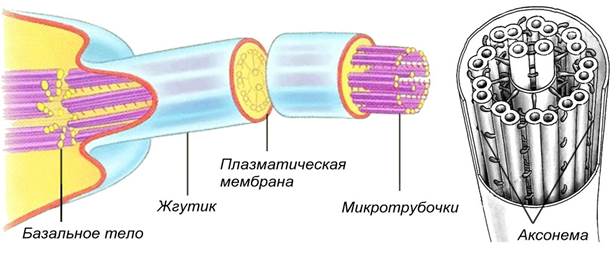
Рис. Строение жгутиков и ресничек эукариот
Включения — временные компоненты цитоплазмы, то возникающие, то исчезающие. Как правило, они содержатся в клетках на определенных этапах жизненного цикла. Специфика включений зависит от специфики соответствующих клеток тканей и органов. Включения встречаются преимущественно в растительных клетках. Они могут возникать в гиалоплазме, различных органеллах, реже в клеточной стенке.
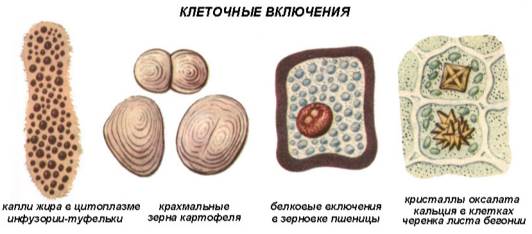
В функциональном отношении включения представляют собой либо временно выведенные из обмена веществ клетки соединения (запасные вещества — крахмальные зерна, липидные капли и отложения белков), либо конечные продукты обмена (кристаллы некоторых веществ).
Крахмальные зерна. Это наиболее распространенные включения растительных клеток. Крахмал запасается у растений исключительно в виде крахмальных зерен. Они образуются только в строме пластид живых клеток. В процессе фотосинтеза в зеленых листьях образуется ассимиляционный, или первичный крахмал. Ассимиляционный крахмал в листьях не накапливается и, быстро гидролизуясь до сахаров, оттекает в части растения, в которых происходит его накопление. Там он вновь превращается в крахмал, который называют вторичным. Вторичный крахмал образуется и непосредственно в клубнях, корневищах, семенах, то есть там, где он откладывается в запас. Тогда его называют запасным. Лейкопласты, накапливающие крахмал, называют амилопластами. Особенно богаты крахмалом семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений.
Липидные капли. Встречаются практически во всех растительных клетках. Наиболее богаты ими семена и плоды. Жирные масла в виде липидных капель — вторая по значению (после крахмала) форма запасных питательных веществ. Семена некоторых растений (подсолнечник, хлопчатник и т.д.) могут накапливать до 40% масла от массы сухого вещества.
Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. Липидные капли могут накапливаться и в лейкопластах, которые называют элайопластами.
Белковые включения образуются в различных органеллах клетки в виде аморфных или кристаллических отложений разнообразной формы и строения. Наиболее часто кристаллы можно встретить в ядре — в нуклеоплазме, иногда в перинуклеарном пространстве, реже в гиалоплазме, строме пластид, в расширениях цистерн ЭПР, матриксе пероксисом и митохондриях. В вакуолях встречаются как кристаллические, так и аморфные белковые включения. В наибольшем количестве кристаллы белка встречаются в запасающих клетках сухих семян в виде так называемых алейроновых3 зерен или белковых телец.
Запасные белки синтезируются рибосомами во время развития семени и откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, и белок кристаллизуется. В результате этого в зрелом сухом семени белковые вакуоли превращаются в белковые тельца (алейроновые зерна).
Включения классифицируют на трофические, представляющие собой запасы питательных веществ (углеводные – гликоген в клетках печени и мышцах, белковые – желточные включения в яйцеклетках, жировые – капли жира в клетках подкожной жировой клетчатки), секреторные продукты жизнедеятельности клеток желез внешней и внутренней секреции (ферменты, гормоны, слизь и др.), пигментные – содержат различные пигменты (меланин, липофусцин, гемоглобин и др.) и экскреторные – продукты обмена веществ, подлежащие удалению из клетки, (кристаллы щавелевой кислоты, щавелевокислого кальция, мочевина)
§
Ядро– важнейший структурный компонент живых клеток эукариот.
Впервые ядро было описано Р. Броуном в 1831 г. Морфологию и функции ядра исследовали Флемминг, Страсбургер, Чистяков, Геккель, Баранецкий, Навашин, Герасимов, Беляев и др. Большинство клеток содержат одно ядро, но встречаются двуядерные (инфузория-туфелька) и многоядерные (скелетные мышцы, печень) клетки. Некоторые высокоспециализированные клетки утрачивают ядра (эритроциты млекопитающих и клетки ситовидных трубок у покрытосеменных).
Ядро представляет собой эластичное тело, отделенное от цитоплазмы ядерной оболочкой. Форма ядра, как правило, круглая, но бывает веретеновидная, нитевидная, сегментированная (лопастная) и др. Впячивания и выпячивания ядерной оболочки значительно увеличивают поверхность ядра, тем самым усиливая связь ядерных и цитоплазматических структур и веществ. Ядро всегда располагается в цитоплазме.
По физическим и химическим свойствам ядро близко к цитоплазме.
Клетка, утратившая ядро, не может дальше существовать. Ядро также не способно к самостоятельному существованию, поэтому можно сказать, что ядро и цитоплазма образуют взаимозависимую систему.

Рис. Схема ультраструктурой организации интерфазного ядра: 1 — ядерная мембрана с порами (2), 3 — плотный хроматин; 4 — рыхлый хроматин; 5 — ядрышко; 6 — интерхроматиновые гранулы; 7 — перихроматиновые гранулы; 8 — перихроматиновые фибриллы; 9 — кариоплазма.
Ядро состоит из ядерной оболочки, ядерного сока, ядрышка и хроматина.
Ядерная оболочка (кариолемма) очень тонкая (300-500 Ао); образована двумя мембранами (наружной и внутренней), между которыми имеется полость – перинуклеарное пространство. Наружная ядерная мембрана покрыта рибосомами, внутренняя мембрана гладкая. Ядерная оболочка пронизана порами (округлые отверстия диметром 200-300 Ао), через которые между ядром и цитоплазмой происходит обмен различными веществами. Также вещества из ядра в цитоплазму и из цитоплазмы в ядро попадают путем отшнуровывания выростов и выпячиваний ядерной оболочки. Кроме того, мелкие молекулы могут диффундировать через ядерную оболочку. В определенных точках ядерная мембрана непосредственно переходит в мембрану эндоплазматической сети, с которой тождественна по своей физико-химической структуре. Несмотря на активный обмен веществ между ядром и цитоплазмой, ядерная оболочка отграничивает ядерное содержимое от цитоплазмы, делая возможным существование особой внутриядерной среды, отличной от окружающей цитоплазмы.
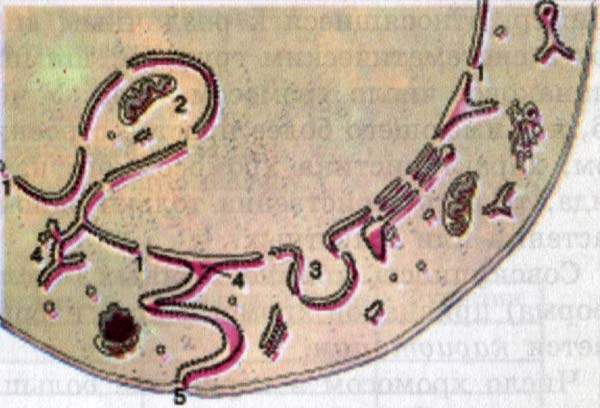
Рис. Пути обмена веществ между ядром и цитоплазмой. 1 — обмен веществ через ядерные поры, 2 — впячивание цитоплазмы внутрь ядра, 3 — впячивание ядерной оболочки, 4 — продвижение ядерной мембраны в эндоплазматическую сеть; 5 — выведение части каналов во внешнее межклеточное пространство.
Ядерный сок (кариоплазма, нуклеоплазма, кариолимфа) представляет собой желеобразный раствор – систему гидрофильных коллоидов – в котором находятся разнообразные белки, нуклеотиды, а также хромосомы и ядрышко. По химическому составу ядерный сок близок к матриксу цитоплазмы, однако в нем значительно выше содержание нуклеотидов. Функция ядерного сока – связь ядерных структур.
Ядрышко образование более плотное, чем основная масса ядра, собственной оболочки не имеет, состоит из крупных гранул, по форме и размерам близко к рибосомам. Матрикс ядрышка имеет жидку консистенцию. Формируется ядрышко в области вторичной перетяжки (ядрышковый организатор). Функция ядрышка – синтез р-РНК и соединение их с белками, т.е. сборка субъединиц рибосом.
Хроматин – глыбки, гранулы и нитчатые структуры, окрашивающиеся некоторыми красителями (гематоксилином, софранином, кармином и др.). С химической точки зрения хроматин – дезоксирибонуклеопротеид (ДНП, комплекс ДНК и белков-гистонов). Гистоны обладают основными (щелочными) свойствами благодаря высокому содержанию в них основных аминокислот. По преобладающему содержанию аминокислот выделяют пять важнейших гистонов:
— гистон Н1 имеет высокое содержание лизина;
— гистон Н2b лизина содержит меньше, чем Н1;
— гистон Н2a имеет высокое содержание лизина и аргинина;
— гистон Н3 содержит большое количество аргинина;
— гистон Н4 богат аргинином и глицином.
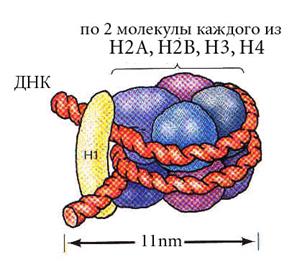
Все гистоны хорошо растворимы в кислых средах. Гистоновые белки с неодинаковой прочностью связываются с ДНК. Поэтому они обладают различной способностью менять пространственное расположение нити ДНК и влиять на участие ДНК в процессе транскрипции. Молекулы гистонов соединяются с ДНК в основном за счет электростатических связей между отрицательно заряженными фосфатными группами молекулы ДНК и положительно заряженными группами гистоновых аминокислот, обладающих щелочными свойствами. В результате образуется нуклеосома. Нуклеосома – это комплекс участка ДНК с гистонами. Он имеет небольшую длину и периодически повторяется по всей длине ДНК. В состав нуклеосомы входит от 160 до 240 нуклеотидных пар и по 2 молекулы каждой фракции гистонов Н2a, Н2b, Н3 и Н4 – всего 8 молекул, соединенных между собой при помощи своих гидрофобных участков. Основной участок нуклеосомы представляет собой цилиндр (октамер) диаметром 11 нм и толщиной 5,7 нм, вокруг которого двойная спираль образует около двух витков и переходит на следующий цилиндр. Длина «накрученного» фрагмента ДНК составляет примерно 60 нм.
Хроматин представляет собой деспирализованные и спирализованные (уплотненные) участки хромосом. Спирализованные участки в генетическом отношении инертны. Передачу наследственной информации осуществляют рыхло спирализованные эухроматиновые участки. В делящихся клетках хромосомы сильно спирализуются, укорачиваются и приобретают компактные размеры и форму.
§
Первый уровеньупаковки ДНК – нуклеосомный. Нуклеосомная нить хроматина (см. выше) имеет диаметр около 13 нм. После упаковки длина молекулы ДНК уменьшается в 5-7 раз. Нуклеосомный уровень обнаруживается в электронном микроскопе в интерфазе и при митозе.
Второй уровень упаковки – соленоидный (супернуклеосомный). Нуклеосомная нить конденсируется, её нуклеосомы «сшиваются» гистоном Н1 и образуется спираль диаметром около 25 нм. Один виток спирали содержит 6-10 нуклеосом. Нить укорачивается еще в 6 раз. Супернуклеосомный уровень упаковки обнаруживается в электронном микроскопе как в интерфазных, так и в митотических хромосомах.
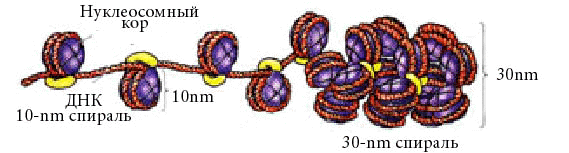 Третий уровень упаковки – хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель и изгибов. Она составляет основу хроматиды и обеспечивает хроматидный уровень упаковки. Он обнаруживается в профазе. Диаметр петель около 50 нм. Нить ДНП укорачивается в 10-20 раз.
Третий уровень упаковки – хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель и изгибов. Она составляет основу хроматиды и обеспечивает хроматидный уровень упаковки. Он обнаруживается в профазе. Диаметр петель около 50 нм. Нить ДНП укорачивается в 10-20 раз.

Четвертый уровеньупаковки – уровень метафазной хромосомы. Хроматиды в заканчивают спирализацию: происходит укорочение в 20 раз. Метафазные хромосомы имеют длину от 0,2 до 150 мкм и диаметр от 0,2 до 5,0 мкм. Общий итог конденсации – укорочение нити ДНП в 10 000 раз!!!


Хромосомы – плотные, интенсивно окрашивающиеся структуры, единицы морфологической организации генетического материала и обеспечивают его точное распределение при делении клетки. Хромосомы лучше всего различимы (и изучаются) на стадии метафазы митоза.
Метафазные хромосомы имеют вид коротких нитевидных фигур, или изогнутых палочек, состоящих из двух продольных нитей ДНП – хроматид. Хроматиды в точке перегиба (первичная перетяжка) соединены центромерой, к которой прикрепляются нити веретена деления. Центромера делит тело хромосомы на два плеча. Участок каждого плеча вблизи центромеры называется проксимальным, удаленный от неё – дистальным. Концевые отделы дистальных участков называются теломерами. Теломеры препятствуют соединению концевых участков хромосом. Потеря этих участков может сопровождаться хромосомными перестройками. Кроме первичной перетяжки, отдельные хромосомы имеют вторичные, не вызывающие перегиба хро
мосомы. Положение вторичной перетяжки, ее длина постоянны для каждого вида хромосом. Некоторые хромосомы имеют еще спутник – округлое или
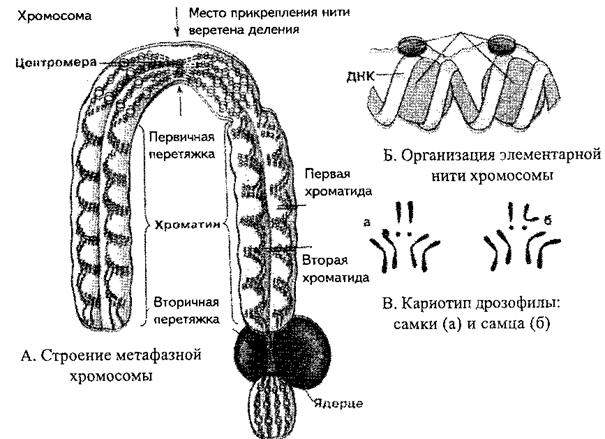
палочковидное тело той же природы. С основным телом хромосомы спутник соединяется тонкой хроматиновой нитью. Иногда спутником считают часть хромосомы, отделенную вторичной перетяжкой. Хромосомы со спутниками характерны для растительных клеток.
Типы хромосом. В зависимости от положения центромеры различают следующие типы хромосом:
— метацентрические (равноплечие), центромера расположена посередине и плечи примерно одинаковой длины (3);
—субметацентрические (неравноплечие), центромера умеренно смещена от середины хромосомы, плечи имеют разную длину (2);
— акроцентрические (палочковидные), центромера значительно смещена к одному концу хромосомы, или располагается в ее теломерном участке, в результате одно плечо очень короткое или отсутствует (1).

Рис. Типы хромосом.
Изучение хромосом позволило установить:
— во всех соматических клетках любого организма число хромосом одинаково;
— в половых клетках содержится всегда вдвое меньше хромосом, чем в соматических клетках данного вида организмов;
— у всех организмов, относящихся к данному виду, число хромосом в клетках одинаково.
В качестве примера ниже приведены диплоидные числа хромосом в ядрах соматических клеток некоторых видов организмов.
Малярийный плазмодий – 2; Лошадиная аскарида – 2; Дрозофила – 8: Вошь головная – 12; Шпинат – 12; Муха домашняя – 12; Окунь – 28; Человек – 46; Ясень – 46; Шимпанзе – 48; Таракан – 48; Перец – 48; Овца – 54; Собака – 78; Голубь – 80; Сазан – 104.
Как видно, число хромосом не зависит от уровня организации и не всегда указывает на филогенетическое родство, поскольку одно и то же число хромосом может встречаться у видов очень далеких в систематическом отношении и сильно отличаться у близких по происхождению организмов. Таким образом, число хромосом не является видоспецифичным признаком. Однако, характеристика хромосомного набора в целом видоспецифична, т.е. свойственна только одному какому-то виду организмов. Совокупность количественных (число) и качественных (форма) признаков хромосомного набора соматической клетки называется кариотипом.
Число хромосом в кариотипе всегда четное. Это объясняется тем, что в соматических клетках всегда находятся две одинаковые по форме и размерам хромосомы: одна происходит от отцовского организма, другая – от материнского. Парные хромосомы, одинаковые по форме и размерам и несущие одинаковые гены, называются гомологичными. Хромосомы из разных пар – называют негомологичными. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, называют двойным, или диплоидным набором(2n). В половые клетки из каждой пары гомологичных хромосом попадает только одна, поэтому хромосомный набор гамет называют одинарным, или гаплоидным набором. Количество ДНК, содержащееся в одинарном наборе хромосом – 1с, соответственно в двойном наборе количество ДНК – 2с. Хромосомы в составе кариотипа делят также на аутосомы, или неполовые, одинаковые у особей мужского и женского пола, и гетерохромосомы, или половые, участвующие в определении пола и различающиеся у самцов и самок. Кариотип человека представлен 46 хромосомами (23 пары): 44 аутосомы и 2 половые хромосомы (у женщины две одинаковые Х-хромосомы, у мужчины Х- и Y-хромосомы).
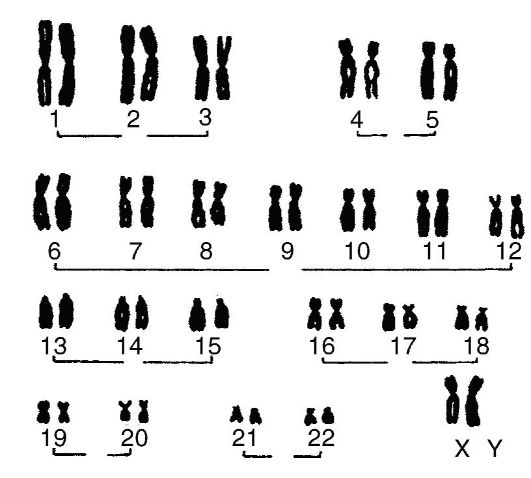
Рис. Кариотип человека.
Правила хромосом.
Правило постоянства числа хромосом: соматические клетки организма каждого вида имеют строго определенное количество хромосом (у человека – 46, у дрозофилы – 8).
Правило парности хромосом: каждая хромосома в диплоидном наборе имеет гомологичную – сходную по размерам, расположению центромеры и содержанию генов.
Правило индивидуальности хромосом: каждая пара хромосом отличается от другой пары размерами, расположением центромеры и содержанием генов.
Правило непрерывности хромосом: в процессе удвоения генетического материала новая молекула ДНК синтезируется на основе информации старой молекулы ДНК (реакция матричного синтеза – каждая хромосома от хромосомы).
Таблица 1
Сравнительная характеристика прокариотической и эукариотической клеток
| ПРОКАРИОТИЧЕСКАЯ КЛЕТКА | ЭУКАРИОТИЧЕСКАЯ КЛЕТКА |
| Цитоплазматическая мембрана | Цитоплазматическая мембрана |
| Клеточная стенка из муреина | Клеточная стенка из целлюлозы (растения) или хитина (грибы) |
| Цитоплазма | Цитоплазма |
| Оформленное ядро отсутствует | Ядро, отграниченное от цитоплазмы ядерной оболочкой |
| Отсутствуют органоиды: митохондрии, комплекс Гольджи, ЭПС, лизосомы, пластиды | Имеются митохондрии, комплекс Гольджи, ЭПС, лизосомы, пластиды. |
| Мезосомы выполняют функции ряда органоидов. | Мезосомы отсутствуют |
| Рибосомы | Рибосомы |
| Генетический аппарат представлен одной кольцевой молекулой ДНК | ДНК линейной структуры в комплексе с гистоновыми белками |
| Набор хромосом гаплоидный | Набор хромосом диплоидный, или у некоторых фаз жизни гаплоидный. |
| Простое бинарное деление | Митоз, мейоз, амитоз, эндомитоз, политения. |
Таблица 2.
Сравнительная характеристика клеток растений и животных
| РАСТИТЕЛЬНАЯ КЛЕТКА | ЖИВОТНАЯ КЛЕТКА |
| Клеточная стенка, состоящая из целлюлозы, или хитина | Клеточной стенки нет |
| Нет клеточного центра | Клеточный центр |
| Крупная центральная вакуоль, содержащая клеточный сок | Вакуоли мелкие (пищеварительные, сократительные) |
| Пластиды | Пластид нет |
| Органические вещества синтезируют из неорганических, используя энергию солнечного света | Органические вещества синтезируются на основе готовых органических веществ, поступающих с пищей. |

Ключевые слова и понятия:
Активный транспорт
Аппарат Гольджи
Аутосомы
Биологическая мембрана
Вакуоль
Включения
Гаплоидный набор хромосом
Гетерохромосомы
Грана
Гомологичные хромосомы
Диплоидный набор хромосом
Диффузия
Жгутик
Кариоплазма
Кариотип
Криста
Лейкопласты
Микротрубочки
Микрофиламенты
Митохондрия
Негомологичные хромосомы
Органоиды
Осмос
Пассивный транспорт
Пиноцитоз
Пропластида
Реснички
Рибосома
Строма
Тилакоид
Фагоцитоз
Хлоропласт
Хроматин
Хромопласт
Хромосома
Центриоль
Центросома
Циклоз
Цитоплазматическая мембрана
Экзоцитоз
Эндоплазматическая сеть агранулярная
Эндоплазматическая сеть гранулярная
Эндоцитоз
Ядро
Ядрышко