

- Количество газов в крови (мл/л)
- Расход кислорода при резке труб. резка труб кислородом.
- § 12. ручная газовая резка труб. расход пропана и кислорода на 1 м реза
- Нормы расчета горючих газов и окислителя
- Определение норматива расхода газов
- Определение значения допустимого расхода и скорости резания
- Гильотина
- § 12. ручная газовая резка труб
Количество газов в крови (мл/л)
| кровь | О2 | СО2 |
| артериальная | 180-200 | |
| венозная | 130-150 |
Кислородная емкость крови (КЕК) – максимальное количество кислорода, которое может быть связано 1 л крови. Составляет 180-200 мл/л (табл. 4).
Кислородную емкость крови определяют по концентрации Нb с двухвалентным железом (Fe2 ). Максимально 1 моль гемоглобина может присоединить 4 моля О2 к своим 4 молям железа гема. Принимая во внимание молекулярную массу (64500 Да) 1 г гемоглобина связывает 4/64500 = 0,062 ммоль О2. Так как молярный объем идеального газа равен 22,4 л/моль, то 1г гемоглобина связывает 1,39 мл О2 (0,062´22,4).
Измерения непосредственно в крови демонстрируют меньшую величину, так как некоторая часть гемоглобина в организме в нормальных условиях находится в измененной форме, которая не может связываться с О2 (например карбоксигемоглобин НbCО, метгемоглобин). Для практических целей применяют величину, называемую числом Гюфнера, приблизительно равную 1,34 мл О2 на 1 г Нb.
Коэффициент утилизации кислорода (КУО2)представляет собой часть кислорода, поглощаемую тканями из капиллярного русла.
КУО2 —это процентное отношение доли кислорода, используемой тканями (разности концентраций кислорода в артериальной и венозной крови), к общему содержанию его в артериальной крови:
КУО2 = VO2а— VO2в / VO2а x 100 %
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 25-35%). Это позволяет тканям приспосабливаться к снижению доставки кислорода
Зависимость степени оксигенации гемоглобина от парциального давления О2 в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой (рис. 15).
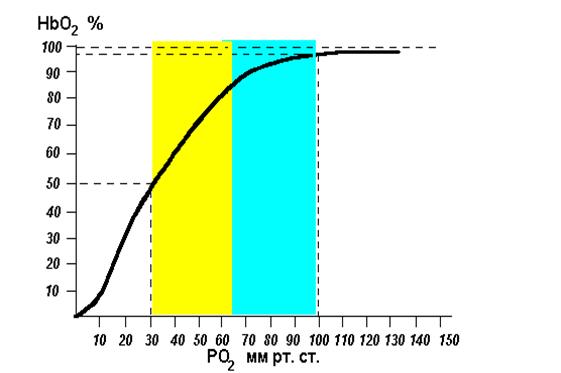
Рис. 15. Кривая диссоциации оксигемоглобина, или сатурационная кривая
Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях. Кривая имеет S-образную форму, что обусловлено кооперативным взаимодействием четырех субъединиц, составляющих тетрамер гемоглобина. Присоединение О2 к гемму одной субъединицы повышает афинность (сродство) для его соединения с остальными субъединицами. Важным параметром для описания этой кривой является парциальное давление О2 при половинном насыщении гемоглобина. В человеческой крови Р0,5=27 мм рт.ст.
S-образная форма кривой связывания гемоглобином О2 имеет большое значение для транспортной функции крови. В области значений выше 65 мм рт.ст. кривая плоская, изменение Ро2 только немного меняет насыщение кислородом. Это область нормальных значений альвеолярного Ро2, которое может немного снижаться без заметного уменьшения насыщения гемоглобина кислородом в крови капилляров легких (например при подъеме в горы). Отвесный спад в нижней части кривой связывания (менее 65 мм рт.ст.) гарантирует, что Ро2 в капиллярной крови периферических тканей, несмотря на отдачу кислорода, остается достаточно высоким, чтобы обеспечить ткани кислородом путем диффузии.
Ряд факторов влияет на афинность (сродство) гемоглобина к О2. При этом в первую очередь изменяется положение кривой и гораздо меньше – ее форма. Различные метаболические факторы вызывают смещения кривой диссоциации влево или вправо (рис. 16).
ВЛЕВО ВПРАВО

Рис. 16. Сдвиги кривой диссоциации оксигемоглобина
Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата.
1) Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2). Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора». Эффект Бора может быть связан также с изменением Рсо2: его повышение сдвигает кривую связывания О2 вправо, понижение — влево. Это влияние основывается, в первую очередь, на том, что повышение Рсо2 ведет к снижению рH и наоборот. Влияние самих молекул СО2 на афинность гемоглобина к О2, напротив, незначительно.
2) Влияние температуры: рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей вызывает сдвиг влево кривой диссоциации оксигемоглобина. Значение влияния температуры для гомойотермных организмов небольшое, хотя температура крови на периферии тела может сильно отличаться от температуры всего организма.
3) Эффект 2,3-дифосфоглицерата:в его отсутствии афинность гемоглобина к кислороду очень высока. Главная функция 2,3-дифосфоглицерата состоит в смещении кривой связывания О2 вправо, в область, которая соответствует физиологическим условиям.
Контрольные вопросы
1. Какие транспортные формы кислорода существуют ?
2. Что такое кислородная емкость крови, как она определяется ?
3. Что такое коэффициент утилизации кислорода, как он определяется ?
4. Почему кривая диссоциации оксигемоглобина (сатурационная кривая) имеет S- образную форму ?
5. Какое биологическое значение крутой и отлогой частей кривой диссоциации оксигемоглобина (сатурационной кривой) ?
6. Какие факторы влияют на афинность (сродство) гемоглобина к О2 ?
§
В венозной крови содержится около 580 мл/л СО2. Транспорт обеспечивается в таких формах, как: 1) растворенный СО2 в плазме крови (5—10%); 2) в виде гидрокарбонатов (80—90%); 3) карбаминовые соединения эритроцитов (5—15%).
Небольшая часть СО2 транспортируется в легкие в растворенном виде (0,3 мл/100 мл крови). Растворенный в крови СО2 реагирует с водой:
CO2 Н2О = H2CO3
В плазме крови эта реакция протекает медленно и не имеет особого значения. Но в эритроцитах имеется цинк-содержащий фермент — карбоангидраза — который смещает равновесие реакции вправо (в сторону образования угольной кислоты). Образование H2CO3 происходит в 1000 раз быстрее, чем в плазме, кроме того, около 99,9% H2CO3 диссоциирует с образованием HCO3—— и иона водорода (H ):
CO2 Н2О =H2CO3 = HCO—3 H
Образующиеся протоны (H ) нейтрализуются гемоглобиновым буфером (H Hb = HHb). Образующийся HCO3— выходит из эритроцитов в плазму, для
поддержания электронейтральности в эритроциты поступают ионы Cl —.
В эритроците CO2 может также связываться гемоглобином с образованием HbCO2. Как и в первом случае, образующийся при этом H связывается гемоглобиновым буфером.
Как сатурация гемоглобина кислородом коррелирует с PО2, так и общее
содержание CO2 в крови коррелирует с PCO2 и описывается кривой диссоциации CO2 (рис. 17).

Рис. 17. Кривая диссоциации двуокиси углерода
Даже при выраженных нарушениях Va/Q (то есть при выраженной легочной патологии) Paco2, как правило, остается в пределах нормальных значений. Это является следствием того, что кривая диссоциации CO2 (рис. 17) нарастает монотонно. Артериовенозная разница по Pco2 в покое обычно составляет 5 мм рт. ст.и редко превышает 10 мм рт. ст. При данном значении Pco2 деоксигенированная кровь содержит большее количество CO2, чем оксигенированная (эффект Холдена). В отличие от кривой насыщения Hb кислородом кривая содержания CO2 не имеет плато и в клинически значимом диапазоне представляет собой прямую линию.
В венозной крови, притекающей к капиллярам легких, напряжение СО2 составляет в среднем 46 мм рт.ст., а в альвеолярном воздухе парциальное давление СО2 равно в среднем 40 мм рт.ст., что обеспечивает диффузию СО2 из плазмы крови в альвеолы легких по концентрационному градиенту.
Эндотелий капилляров проницаем только для молекулярного СО2 как полярной молекулы. Из крови в альвеолы диффундирует физически растворенный в плазме крови молекулярный СО2. Кроме того, в альвеолы легких диффундирует СО2, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах. Молекулярный СО2 проходит аэрогематический барьер, а затем поступает в альвеолы. В норме через 1 с происходит выравнивание концентраций СО2 на альвеолярно-капиллярной мембране, поэтому за половину времени капиллярного кровотока происходит полный обмен СО2 через аэрогематический барьер. Реально равновесие наступает несколько медленнее. Это связано с тем, что перенос СО2, так же как и О2, ограничивается скоростью перфузии капилляров легких.
Контрольные вопросы
1. Какие транспортные формы углекислого газа существуют ?
2. Какая форма транспорта углекислого газа основная ?
3. Почему кривая диссоциации двуокиси углерода в виде прямой линии ?
3.3. Тестовые задания и ситуационная задача
Выберите один правильный ответ.
20. УГЛЕКИСЛЫЙ ГАЗ В КРОВИ ТРАНСПОРТИРУЕТСЯ В ВИДЕ
1) растворенного газа, связанного с гемоглобином, в составе бикарбонатов
2) только в растворенном виде
3) только в составе бикарбонатов
21. ДИСССОЦИАЦИЯ ОКСИГЕМОГЛОБИНА УВЕЛИЧИВАЕТСЯ ПРИ УСЛОВИИ
1) увеличения pH крови, уменьшения содержания CO2 в крови
2) уменьшения температуры тела, увеличения содержания карбоксигемоглобина
3) увеличения pH крови, повышения температуры тела, увеличения содержания CO2 в крови
4) увеличения pH крови и повышения температуры тела
22. КИСЛОРОДНАЯ ЕМКОСТЬ КРОВИ – ЭТО
1) максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом
2) диффузия кислорода и альвеолярного воздуха в кровь
3) часть кислорода, поглощаемого тканями из венозной крови
4) часть кислорода, поглощаемого тканями из артериальной крови
23. КОЭФФИЦИЕНТОМ УТИЛИЗАЦИИ КИСЛОРОДА НАЗЫВАЮТ
1) максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом
2) большую часть кислорода, содержащуюся в артериальной крови
3) часть кислорода, поглощённую тканями из венозной крови
4) часть кислорода, поглощённую тканями из артериальной крови
Ситуационная задача 3
У больного резко нарушен транспорт гемоглобина. Какое терапевтическое воздействие может помочь в обеспечении тканей кислородом ?
Газообмен в тканях
Обмен СО2 между клетками тканей с кровью тканевых капилляров определяется следующими механизмами. В процессах окисления в тканях образуется СО2. Обычно в большинстве тканей рСО2 близок в 50-60 мм рт. ст. В крови, поступающей в артериальный конец капилляров раСО2= 40 мм рт.ст. Наличие градиента заставляет СО2 диффундировать из тканевой жидкости к капиллярам. Интенсивность окисления в различных тканях не одинакова. Поэтому в смешанной венозной крови, поступающей в правое предсердие в покое, рvcо2 равно 46 мм рт.ст. Кровь, проходящая через легкие отдает не весь СО2. Большая часть его сохраняется в артериальной крови.
В ходе газообмена СО2 между тканями и кровью содержание НСО3— в эритроците повышается и они начинают диффундировать в кровь (рис. 18). Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1—. Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов. В условиях in vitro образование молекулярного СО2 из гидрокарбонатов происходит чрезвычайно медленно и диффузия этого газа занимает около 5 мин, тогда как в капиллярах легкого равновесие наступает через 1 с. Это определяется функцией фермента карбоангидразы угольной кислоты. В функции карбоангидразы выделяют следующие типы реакций:
СО2 Н2О =H2СО—3 = H НСО3—

Рис. 18. Газообмен в тканях
Карбаминовый комплекс СО2 с гемоглобином образуется в результате реакции СО2 с радикалом NH2 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н ; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2.
Контрольные вопросы
1. Чему равно напряжение углекислого газа в тканях, артериальной и венозной крови ?
2. Какова последовательность газообмена в тканях ?
3. Какой тип реакции обеспечивает карбоангидраза ?
4.1. Тестовые задания
Выберите один правильный ответ.
24. ФЕРМЕНТ КАРБОАНГИДРАЗА НАХОДИТСЯ
1) в слизи
2) в сурфактанте
3) в эритроцитах
4) в плазме крови
25. НАПРЯЖЕНИЕ УГЛЕКИСЛОГО ГАЗА В ВЕНОЗНОЙ КРОВИ СОСТАВЛЯЕТ
1) 100 мм рт.ст.
2) 46 мм рт.ст.
3) 40 мм рт.ст.
4) 15 мм рт.ст.
Тканевое дыхание
Более 90% всей энергии в организме образуется в результате горения водорода. Сам данный процесс суммарно можно представить в ходе простой химической реакции:
 2Н2 О2 2Н2О
2Н2 О2 2Н2О
Термин «горение» применительно к организму применим весьма условно, хотя суть процесса реакция отображает правильно. Реакция взаимодействия водорода с кислородом с химической точки зрения является окислительно-восстановительной, поэтому применительно к организму используется другой термин — биологическое окисление. Водород используется как главное топливо для образования энергии. В митохондриях поток электронов от водорода устремляется к их главному акцептору кислороду. При этом образуются молекулы воды, которая в энергетической шкале биологических веществ занимает низшую ступеньку и является конечным продуктом тканевого дыхания. Предыдущие этапы дыхания обеспечивают лишь доставку кислорода к клеткам, где в митохондриях функционирует цепь переноса электронов (дыхательная цепь). При этом происходит перенос по цепи переносчиков вначале водорода, а затем электронов на кислород, с постепенным освобождением энергии, значительная часть которой аккумулируется в макроэргических связях АТФ. У теплокровных животных, в том числе и у человека, работа дыхательной цепи одновременно обеспечивает выработку тепла, которое идет на поддержание температуры тела.
Процесс тканевого дыхания носит достаточно сложный и многоступенчатый характер, начало изучения, которого было положено в XVIII веке. Следует отметить, что работы ученых, о вкладе которых в изучения процессов биологического окисления и тканевого дыхания будет говориться ниже, не потеряли значения до настоящего времени.
§
Первые представления о тканевом дыхании связаны с Лавуазье, который один из первых указал на то, что жизнь поддерживается кислородом. Он назвал дыхание процессом биологического «горения», подобным горению угля, только очень медленным. Таким образом, Лавуазье были обнаружены сходство и отличия между процессами горения и биологического окисления, которые заключались в следующем:
Сходства:
1. Как горение, так и биологическое окисление идут с потреблением кислорода;
2. Конечными продуктами горения и биологического окисления являются вода и углекислый газ.
Отличия биологического окисления от горения заключаются в следующем:
1. Протекает при температуре человеческого тела;
2. Протекает в водной среде;
3. Отсутствует пламя.
Другой вопрос, который долгое время занимал ученых, был о происхождении энергии активации необходимой для взаимодействия водорода и кислорода. Дело в том, что в молекулярной форме кислород относительно малоактивен, поэтому «гремучую смесь» (смесь кислорода с водородом) необходимо нагреть, а водород выходящий из трубки генератора — поджечь. В условиях организма это невозможно, поэтому дискуссия о происхождении энергии активации в процессах биологического окисления получила дальнейшее развитие. В 1897 году была обоснована первая гипотеза тканевого дыхания, названная гипотезой перекисного (пероксидного) окисления. Ее разрабатывали независимо А.Н. Бах в России и Энглер в Германии. Суть данной гипотезы состоит в том, что при дыхании, как считали авторы, происходит активирование молекул О2 за счет энергии самоокисляющихся веществ, образование пероксидов и разложение их с участием другого вещества:
 1. О = О – О – О – (активный кислород)
1. О = О – О – О – (активный кислород)

О
 2. – О – О – А (субстрат) А (пероксид)
2. – О – О – А (субстрат) А (пероксид)
О

О
 3. А В (второй субстрат) АО ВО
3. А В (второй субстрат) АО ВО
О
Для указанных целей необходимо последовательное действие двух ферментов — оксигеназы и пероксидазы. Впоследствии оказалось, что это не главный, а частный случай окисления веществ при дыхании. В настоящее время известно, что это тип медленного окисления органических веществ имеет место в микросомах печени, а не в митохондриях.
Идея активирования кислорода как основного механизма тканевого дыхания разрабатывалась известным немецким ученым Варбургом, создавшим первые аппараты для изучения тканевого дыхания. Он считал, что активирование кислорода есть ключевой процесс в тканевом дыхании, в результате чего кислород соединяется с водородом и образуется вода.
В 1912 г. Варбург открыл гемосодержащий протеид, названный в последствии цитохромоксидазой, которая активирует кислород. Однако, после открытия в том же году Бателли и Штерном дегидрогеназ ученых захватила идея активирования не кислорода, а водорода субстрата как основного звена тканевого дыхания. В.И. Палладин (1912) предложил схему дыхания, по которой дегидрирование является важнейшим звеном дыхания:


А · Н
2
(субстрат) ½ О
2
Н
2
О
Вскоре Виланд и Тунберг доказали, что возможно активирование водорода субстрата с помощью дегидрогеназы. Гипотеза Палладина получила подтверждение. Примерить дегидрогеназную концепцию Палладина и оксидазную Варбурга в тканевом дыхании удалось после открытия в 1933 г. Кейлином цитохромов, являющихся промежуточными переносчиками электронов от водорода к кислороду. На самом деле Кейлин переоткрыл цитохромы, которые были впервые описаны Мак-Мунном в 1886 г. и названные им гистогематинами.
По современным представлениям, в реально работающей дыхательной цепи присутствуют как элементы активации водорода, так и активации кислорода.
Активация водорода достигается взаимодействием его с коферментами дегидрогеназ. Так редокс-потенциал (о природе и величине редокс-потенциала будет сказано ниже) газообразного водорода равен 0 (нулю), а когда водород в составе кофермента НАДН·Н редокс-потенциал снижается до величины -0,32 В, что говорит об резком увеличении восстанавливающих свойств водорода.
Активация кислорода достигается путем взаимодействия на уровне цитохромоксидазы двух электронов, идущих по дыхательной цепи с молекулой О2. В результате неполного восстановления молекулы О2 (для полного восстановления молекулы О2 необходимо 4 электрона) образуется активная форма кислорода — пероксидный свободный радикал. Таким образом, теория активации кислорода также нашла свое подтверждение.
Контрольные вопросы
1. Что такое тканевое дыхание ?
2. В чем сходство и отличие между процессами горения и биологического окисления в организме ?
3. В чем суть гипотез А.Н. Баха и В.И. Палладина ?
§
Дыхательной цепи
В переносе электронов от субстрата к молекулярному кислороду принимают участие:
1. Коферменты на основе витамина В5 (РР) — пиридинзависимые дегидрогеназы, для которых коферментами служат либо НАД, либо НАДФ;
2. Коферменты на основе витамина В2 — флавиновые дегидрогеназы (флавиновые ферменты), у которых роль простетической группы играют ФАД или ФМН;
3. Убихинон (коэнзим Q);
4. Цитохромы, содержащие в качестве простетической группы железопорфириновую кольцевую систему. Среди компонентов дыхательной цепи обнаружены также и железосерные белки (FeS), содержащие негеминовое железо.
5.2.1. Характеристика коферментов на основе витамина В5 (РР)
Витамин B5 является компонентом коферментов. Функции витамина в организме человека выполняют никотиновая кислота и ее амид. Ниацин в организме используется для образования никотинамидных коферментов. Их представителями являются никотинамид-аденин-динуклеотид (НАД , NAD ) и никотинамид-аденин-динуклеотид-фосфат (НАДФ , NADP ). Эти коферменты образуют специфические, каталитически активные комплексы с многочисленными (более 200) дегидрогеназами. Холоферменты участвуют в осуществлении окислительно-восстановительных реакций метаболизма.
В частности, эти комплексы:
1. Облегчают реакции окисления одного субстрата за счет восстановления другого. Эти реакции особенно важны для осуществления окислительных процессов в отсутствии кислорода. Примером подобных реакций может служить обратимое окисление молочной кислоты до пировиноградной, которую катализирует фермент лактатдегидрогеназа (ЛДГ):

 НООC-CН(ОН)-CH3
НООC-CН(ОН)-CH3 НООC-C(О)-CH3
НООC-C(О)-CH3
молочная к-та НАД НАДН∙Н ПВК
Схема 1. Участие кофермента НАД в обратимом окислении молочной кислоты в составе фермента лактатдегидрогеназы
2. Обеспечивают функцию компонентов дыхательной цепи, транспортируя электроны (водород) от субстрата на кислород.
Схема 2. Химическое строение никотинамида

Схема 3. Химическое строение никотиновой кислоты
Помимо окислительно-восстановительных реакций, лежащих в основе энергетического обмена, ниацин участвует в ряде других, не окислительно-восстановительных реакций. В частности доказано, что эти реакции сопровождают передачу информации от клетки к клетке, транскрипцию ДНК, синтез хроматина, дифференциацию клеток и многие другие жизненные процессы. Поступает с пищей, синтезируется в организме из триптофана. Участвует в процессах тканевого дыхания, углеводном, белковом и липидном обмене, оказывает нормализующее влияние на уровень холестерина. Обладает выраженным свойством расширять периферические сосуды

Схема 4. Химическая структура кофермента НАД В рамке показана восстановленная форма активного центра кофермента (НАДН·Н ).
Стрелкой показано расположение фосфатной группы в молекуле НАДФ.
Суточная потребность в витамине В5 (РР) – 15-25 мг. Повышается при физической нагрузке, лактации, низкой температуре, в высокогорье, а также при инфекционных заболеваниях, поражениях желудочно-кишечного тракта и нарушении всасывания. В организме возможен синтез никотиновой кислоты из аминокислоты триптофана. При этом из одной из 50 молекул триптофана образуется одна молекула никотиновой кислоты. Недостаток триптофана в пище резко усугубляет симптомы гиповитаминоза никотиновой кислоты при его наличии.
Контрольные вопросы
1. В какие реакциях в организме принимает участие ниацин ?
2. Сколько составляет суточная потребность в витамине В5?
5.2.2. Характеристика коферментов на основе витамина В2
Относится к группе животных пигментов – флавонов. Рибофлавин хорошо растворим в воде, устойчив в кислых растворах, но легко разрушается в нейтральных и щелочных растворах. Весьма чувствителен к УФ-излучению и сравнительно легко подвергается обратимому восстановлению, присоединяя водород по месту двойных связей и превращается в бесцветную лейкоформу. Это свойство рибофлавина легко окисляться и восстанавливаться лежит в основе его биологического действия в клеточном метаболизме. Участвует в окислительно-восстановительных реакциях, влияя на обмен веществ всего организма.
Витамин B2 интенсифицирует процессы обмена веществ в организме, участвуя в метаболизме белков, жиров и углеводов. Рибофлавин необходим для образования красных кровяных телец и антител, для дыхания клеток и роста. Он облегчает поглощение кислорода клетками кожи, ногтей и волос. Он улучшает состояние органа зрения, принимая, наряду с витамином A, участие в процессах темновой адаптации, снижает усталость глаз и играет большую роль в предотвращении катаракты. Витамин B2 оказывает положительное воздействие на слизистые оболочки пищеварительного тракта. Рибофлавин входит в состав флавиновых коферментов, в частности ФМН (флавиномононуклеотид) и ФАД (флавиноадениндинуклеотид).

6,7-Диметил-9-(D-1-рибитил)-изоаллоксазин
Схема 5. Химическое строение рибофлавина
Различают 2 типа реакций с участием этих коферментов:
1. Фермент осуществляет прямое окисление с участием кислорода, т.е. дегидрирование (отщепление электронов и протонов) исходного субстрата или промежуточного метаболита. К ферментам этой группы относятся оксидазы L- и D-аминокислот, ацил-КоА-дегидрогеназа, глициноксидаза и др.


Схема 6. Химическое строение окисленной формы кофермента ФАД (ФМН). (цифрами 1 и 5 показаны места присоединения водорода в окислительно-восстановительных реакциях с образованием восстановленной формы кофермента ФАДН2)
В качестве примера можно привести следующую схему окисления с участием ФАД:
(суб. восс.ф.) (суб. окис.ф.)

 R-CH2-CH2-COOH R-CH=CH-COOH
R-CH2-CH2-COOH R-CH=CH-COOH
ФАД ФАДН2
Схема 7. Участие кофермента ФАД в окислении карбоновых кислот
2. Характеризуется переносом электронов и протонов в системе биологического окисления.
ФМН и ФАД прочно связываются с белковым компонентом молекулы фермента, иногда ковалентно, как в молекуле сукцинатдегидрогеназы. В составе дыхательной цепи ФМН в качестве кофермента входит в состав НАД·Н-дегидрогеназы, которая акцептирует водород, отщепляемый от НАДН·Н . Молекулярная масса НАД·Н-дегидрогеназы около 106. Этот флавиновый фермент тесно связан с железосерным белком, участвующим передаче электронов на коэнзим Q. НАД·Н-дегидрогеназа пересекает поперек внутреннюю мембрану митохондрий, находясь в окружении липидов. Активный центр ее обращен к внутренней поверхности этой мембраны, т.е. к матриксу.
Суточная потребность в витамине В2– 2-3 мг. Возрастает при физической нагрузке, лактации, высокой калорийности пищи с большим содержанием белка, при употреблении больших количеств никотиновой кислоты.
Контрольные вопросы
1. Какие функции в организме выполняет рибофлавин ?
2. В состав каких коферментов входит рибофлавин ?
3. Какие реакции в организме протекают с участием этих коферментов?
4. Сколько составляет суточная потребность в витамине В2?
§
Кофермент Q (КоQ) был впервые описан Мортоном, который показал, что данное соединение имеет повсеместное распространение в клетках и назвал его убихиноном. Позже было установлено, что убихинон, подобно НАД и ФАД, может играть роль промежуточного переносчика водородных атомов, т.е. электронов и протонов.

Схема 8. Строение и участие убихинона (кофермента Q) в окислительно-восстановительных реакциях
КоQ является производным бензохинона с длинной боковой цепью, включающей у различных организмов от 6 до 10 изопреновых звеньев. Человеческий убихинон содержит 10 изопреновых единиц (КоQ10). Наличие изопренового «хвоста» придает молекуле КоQ хорошую растворимость в липидах, поэтому относительно небольшая молекула убихинона легко перемещается от наружного к внутреннему краю внутренней мембраны митохондрий и наоборот.
Считают, что восстановленная форма флавиновых ферментов в цепи дыхательных катализаторов передает атомы водорода (протоны и электроны) КоQ. Как всякий хинон, КоQ может существовать в окисленной и восстановленной форме, что и обеспечивает его функционирование в дыхательной цепи.
Убихинон синтезируется в тканях человека. Источником его образования служат мевалоновая кислота и продукты обмена фенилаланина и тирозина. Признаки недостаточности КоQ у человека не описаны, но при некоторых состояниях повышается потребность в нем. На этом основании его стали относить к витаминоподобным веществам. Потребность в убихиноне для человека неизвестна.
Контрольные вопросы
1. Что является источником образования убихинона ?
2. Какое строение имеет убихинон (кофермент Q) ?
Строение и характеристика цитохромов и железосерных белков
Дальнейший перенос электронов от КоQ (или восстановленной формы флавинового фермента) на кислород осуществляет система цитохромов. В процессе тканевого дыхания наиболее важную роль играют цитохромы b, c1, c, aa3. Все они имеют простетическую группу геминовую группу, близкую к гемму гемоглобина (простетическая группа цитохрома b, как считают, идентична гему).

Схема 9. Строение гема, входящего в состав цитохромов b и с (цитохромы группы а содержат гем A, отличающийся от остальных набором заместителей в порфириновой системе)
Цитохром b имеет различные формы. По мнению ряда авторов в дыхательной цепи функционируют цитохромы b562 и b566, называемые так по максимуму поглощения света. Они образуют комплекс, пересекающий липидную часть мембраны от внутренней (матриксной) к наружной поверхности.
Цитохром c1 расположен в липидном слое ближе к наружной поверхности внутренней мембраны. Имеет молекулярную массу 40000 Д.
Цитохром c относительно легко переходит в водный раствор. Находится на наружной поверхности внутренней мембраны и, очевидно, может выходить в межмембранное пространство. Молекулярная масса цитохрома c 12000 Д
Цитохромы a и a3 образуют комплекс, называемый цитохромоксидазой. Этот комплекс пересекает мембрану поперек внешней стороны, где в липидном слое находится цитохром а, до внутренней мембраны, где находится цитохром а3. Активный центр цитохрома а3 обращен в матрикс. Молекулярная масса цитохромоксидазы около 450000 Д. В отличии от других цитохромов цитохромоксидаза содержит также Cu 2 .
Все цитохромы, будучи гемопротеидами, при переносе электронов подвергаются обратимому окислению-восстановлению путем белок-белковых взаимодействий. При обратимом окислении меняется степень окисления атома железа от Fe 3 до Fe 2 .
Железосерные белки (FeS) участвуют в переносе электронов и протонов в дыхательной цепи минимум в 2-х участках. Железосерные белки имеют небольшую молекулярную массу порядка 10000 Д. Они содержат негеминовое железо и серу. Семейство этих белков отличается разными окислительными свойствами и значениями редокс-потенциала. Железосерные белки находятся в липидном слое мембраны.
Контрольные вопросы
1. Какие существуют разновидности цитохромов ?
2. В чем отличие различных цитохромов ?
Биологическое окисление
Реакции биологического окисления катализируются ферментами. Об окислении можно говорить в 3-х случаях:
1. Отщепление водорода от окисляемого субстрата;
2. Потеря электронов (например, Fe 2 меняет валентность на Fe 3 );
3. Присоединение к субстрату кислорода.
Все три типа реакций имеют место быть в живой клетке.
Процесс окисления непременно сопряжен с процессом восстановления. Оба вещества — окисляемое и восстанавливаемое образуют окислительно-восстановительную пару или редокс-пару.
Для того чтобы численно отобразить окислительно-восстановительные свойства редокс-пары, введено понятие редокс-потенциал, которые приведены в таблице 5.
Таблица 5
§
| Компонент | Молекулярная масса | Число субъединиц | Простетические группы |
| НАД·Н-дегидрогеназа (комплекс I) | 1 ФМН, 16-24 FeS | ||
| Сукцинатдегирогеназа (комплекс II) | 1 ФАД, 1 гем, 8 FeS | ||
| Убихинон | — | — | |
| Убихинолдегидрогеназа (QH2-дегидрогеназа) (комплекс III) | 6-8 | 3 гем, 2 FeS | |
| Цитохром с | — | 1 гем | |
| Цитохромоксидаза (комплекс IV) | 2 гем, 2 Cu 2 | ||
| Н -АТФ-синтетаза | 8-10 | — |
Как уже было сказано компоненты цепи переноса электронов расположены по градиенту редокс-потенциала. Там где перепад редокс-потенциала между двумя окислительно-восстановительными парами составляет 0,2 В и более, дыхание сопрягается с фосфорилированием и из АДФ и неорганического фосфата синтезируется молекула АТФ. При работе полной дыхательной цепи имеются три пункта сопряжения дыхания и фосфорилирования: 1-й — между НАД·Н-дегидрогеназой и КоQ; 2-й — между цитохромами b и c и 3-й — между цитохромами а и а3. При этом образуются из АДФ и неорганического фосфата 3 молекулы АТФ по уравнению:
 АДФ Н3РО4 АТФ Н2О
АДФ Н3РО4 АТФ Н2О
Следует обратить внимание, что на стадии образования КоQ-Н2 сливаются два потока атомов водорода, вводимых в дыхательную цепь НАД-зависимыми и ФАД-зависимыми дегидрогеназами.
Затем в дыхательной цепи пути электронов и протонов расходятся. Перенос электронов осуществляется с помощью цитохромов. Атом железа в геме может менять валентность, присоединяя или отдавая электрон:
 Fe 3 e— Fe 2
Fe 3 e— Fe 2
 Fe 2 — e— Fe 3
Fe 2 — e— Fe 3
Комплекс цитохромов b и c функционирует как QH2-дегидрогеназа, т.е. осуществляет перенос электронов с QH2 на цитохром с:
 QH2 2с (Fe 3 ) Q 2H 2c (Fe 2 )
QH2 2с (Fe 3 ) Q 2H 2c (Fe 2 )
Электроны последовательно проходят через атомы железа цитохромов b и с1, а затем поступают на цитохром с; протоны при этом освобождаются в раствор. Стехиометрический коэффициент 2 перед символом цитохрома обусловлен тем, что с КоQ-H2 передаются два электрона, а цитохромы за один цикл переносят по одному электрону.
Комплекс цитохромов а и а3 действует как цитохромоксидаза (цитохром-с-оксидаза). Цитохромоксидаза, помимо гема, содержит ионы меди, которые тоже участвуют в переносе электронов меняя валентность:
 Cu 2 e— Cu
Cu 2 e— Cu
 Cu — e— Cu 2
Cu — e— Cu 2
Это комплекс цитохромов переносит электроны с цитохрома с на кислород:
 2с (Fe 2 ) ½ О2 2c (Fe 3 ) O2-
2с (Fe 2 ) ½ О2 2c (Fe 3 ) O2-
Электроны последовательно присоединяются к ионам железа цитохромов а и а3, затем к иону меди и, наконец, попадают на кислород.
Кислород, поступающий в митохондрии из крови, связывается с атомом железа в геме цитохрома а3 в форме молекулы О2 (подобно тому, как он связывается с гемоглобином). Затем каждый из атомов молекулы О2 последовательно присоединяет по два электрона и по два протона, превращаясь в молекулу воды:
 О2 4e— 4Н 2Н2О
О2 4e— 4Н 2Н2О
Укороченная дыхательная цепь предназначена для субстратов, которые не могут окисляться через НАД-зависимые дегидрогеназы, вследствие того, что их редокс-потенциал больше чем у окислительно-восстановительной пары НАДН·Н / НАД . Например, редокс-потенциал у янтарной кислоты (сукцината) составляет -0,01 В. Подобные субстраты окисляются ФАД-зависимыми дегирогеназами. Образующийся при этом ФАДН2 передает водород непосредственно на КоQ, с которого он далее передается на цепь цитохромов. Поскольку в этом случаи в работе не участвует I комплекс, то в ходе дыхательной цепи образуется не 3, а 2 молекулы АТФ. Следует также отметить, что при работе укороченной дыхательной цепи эти 2 молекулы АТФ образуются быстрее, чем 3 при работе полной цепи. В некоторых случаях это бывает важно, когда энергию требуется получить быстро.
Таким образом, через дыхательную цепь атомы водорода пищевых веществ достигают конечного акцептора — атмосферного кислорода. В организме человека в результате тканевого дыхания образуется 300-400 мл воды за сутки (метаболическая вода).
Контрольные вопросы
1. Каким принципам подчиняется работа дыхательной цепи ?
2. Какова структура митохондриальной дыхательной цепи ?
3. Какие существуют основные компоненты цепи переноса электронов ?
4. Как работает полная и укороченная дыхательная цепь ?
Дыхательный контроль
Зависимость дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Суть процесса сводится к тому, что АДФ активирует работу дыхательной цепи, а АТФ ингибирует. Этот механизм регуляции имеет очень важное значение, так в результате его действия скорость синтеза АТФ определяется потребностью клетки в энергии: при увеличении расходования АТФ в клеточных процессах, увеличивается концентрация АДФ, что автоматически приводит к ускорению дыхания и фосфорилирования. Можно сказать, что темп работы митохондриям задается фактическими затратами АТФ.
Механизм дыхательного контроля отличается высокой чувствительностью и точностью, поэтому относительные концентрации АТФ и АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой (т.е. частота оборотов цикла АДФ-АТФ) может изменяться в десятки раз.
Контрольные вопросы
1. Что такое дыхательный контроль?
2. В чем заключается механизм дыхательного контроля ?
§
Ингибиторы тканевого дыхания блокируют один из трех пунктов сопряжения дыхания и фосфорилирования. Все ингибиторы можно разделить на три группы, действие которых представлено на схеме 11.
I группа ингибирует дыхательную цепь на уровне 1-го пункта сопряжения дыхания и фосфорилирования. К данным веществам относятся, например, ротенон («рыбий яд»), амобарбитал (препарат барбитурового ряда). При этом прерывается поступление водорода от субстратов окисляющихся через НАД-зависимые дегидрогеназы. Выключается также первое из трех звеньев фосфорилирования. Данная группа ингибиторов не влияет на использование субстратов, окисляющихся через ФАД-зависимые дегидрогеназы.
 Ротенон ФАДН2 Антимицин А
Ротенон ФАДН2 Антимицин А


 Амобарбитал Карбо- – БАЛ
Амобарбитал Карбо- – БАЛ
 _ ксин
_ ксин



 НАДН-дегидро- QH2 QH2-дегидрогеназа
НАДН-дегидро- QH2 QH2-дегидрогеназа
 геназа ФМН в с
геназа ФМН в с
 H2S, CO, CN–
H2S, CO, CN–
 –
–

 цитохром С цитохромоксидаза ½О2
цитохром С цитохромоксидаза ½О2
 а а3
а а3
Схема 11. Ингибирование ферментов цепи переноса электронов
II группа ингибиторов, к которым относится антимицин А (антибиотик, выделенный из стрептомицетов) блокирует дыхательную цепь на уровне 2-го сопряжения дыхания и фосфорилирования и выключает участок цепи до блока.

Схема 12. Функционирования окислительно-восстановительной пары с участием аскорбиновой и дегидроаскорбиновой кислот
Дыхание возможно только в присутствии аскорбиновой кислоты, которая, превращаясь в дегидроаскорбиновую кислоту по уравнению, представленному ниже, передает водород непосредственно на цитохром с. При этом образуется только одна молекула АТФ на уровне цитохромоксидазы.
III группа ингибиторов тканевого дыхания, к которым относятся цианиды (например, NaCN, KCN), сероводород (H2S), оксид углерода (II), (СО, угарный газ) необратимо блокируют цитохромоксидазу, т.е. 3-й пункт сопряжения дыхания и фосфорилирования.
Это приводит к полному прекращению тканевого дыхания и быстрой гибели клетки от дефицита энергии. Поэтому ингибиторы цитохромоксидазы являются сильнейшими ядами. Оксид углерода (II), кроме того, нарушает транспорт кислорода кровью путем образования с гемоглобином устойчивого соединения — карбоксигемоглобин.
Контрольные вопросы
1. На какие группы по механизму действия классифицируют ингибиторы ?
2. Какие ингибиторы относятся к I группе и как они действуют на цепь переноса электронов ?
3. Какие ингибиторы относятся ко II группе и как они действуют на цепь переноса электронов ?
4. Какие ингибиторы относятся к III группе и как они действуют на цепь переноса электронов ?
5.8. Тестовые задания и ситуационная задача
Выберите один правильный ответ.
26. ЦИАНИДЫ, УГАРНЫЙ ГАЗ, СЕРОВОДОРОД ЯВЛЯЮТСЯ ИНГИБИТОРАМИ ЦИТОХРОМОКСИДАЗЫ
1) аллостерического
2) конкурентного
3) необратимого действия
27. ДОНОРОМ ЭЛЕКТРОНОВ ДЛЯ УКОРОЧЕННОЙ ДЫХАТЕЛЬНОЙ ЦЕПИ ЯВЛЯЕТСЯ
1) пируват
2) малат
3) цитрат
4) сукцинат
5) б-кетоглутатат
28. ПРОСТЕТИЧЕСКОЙ ГРУППОЙ ПЕРВИЧНЫХ АКЦЕПТОРОВ ВОДОРОДА ФЛАВИНОВЫХ ДЕГИДРОГЕНАЗ ЯВЛЯЕТСЯ
1) НАДФ
2) ФАД
3) ФМН
29. В СОСТАВ ПРОСТЕТИЧЕСКОЙ ГРУППЫ НАДН – ДЕГИДРОГЕНАЗНОГО КОМПЛЕКСА ВХОДИТ
1) ФМН
2) ФАД
3) хинон
30. В СОСТАВ ПРОСТЕТИЧЕСКИХ ГРУПП ФЛАВИНОВЫХ ДЕГИДРОГЕНАЗ ВХОДИТ ВИТАМИН
1) В1
2) В2
3) В5
4) В3
5) В6
31. КОЛИЧЕСТВО ЭНЕРГИИ, ВЫДЕЛЯЮЩЕЙСЯ ПРИ ПЕРЕНОСЕ ЭЛЕКТРОНОВ ОТ ФАДН2 К МОЛЕКУЛЯРНОМУ КИСЛОРОДУ, ОБЕСПЕЧИВАЕТ СИНТЕЗ АТФ
1) 3
2) 2
3) 1
32. ПРИ БЛОКИРОВАНИИ АМОБАРБИТАЛОМ ПЕРВОГО ПУНКТА ЦЕПИ ПЕРЕНОСА ЭЛЕКТРОНОВ СИНТЕЗ АТФ В МИТОХОНДРИЯХ ВОЗМОЖЕН ПРИ ИСПОЛЬЗОВАНИИ В КАЧЕСТВЕ СУБСТРАТА
1) малата
2) сукцината
3) б-кетоглутарата
4) глутамата
33. ДЫХАТЕЛЬНЫМ КОНТРОЛЕМ НАЗЫВАЕТСЯ РЕГУЛЯЦИЯ СКОРОСТИ ДЫХАНИЯ
1) цитохромоксидазой
2) НАДН-дегидрогеназой
3) концентрацией АДФ
Ситуационная задача 4
В суспензию митохондрий добавили 0,6 ммоль/л малата и 0,3 ммоль/л АДФ. Поглощение кислорода, начавшееся сразу после добавления малата и АДФ, через некоторое время прекратилось. Почему? Затем к пробе добавили раствор фермента гексокиназы и раствор глюкозы. Поглощение кислорода вновь началось. Почему?
Ситуационная задача 5
Сколько молекул АТФ образуется при окислении сукцината? Как будет влиять амобарбитал на окисление этого соединения?
Регуляция дыхания
Дыхание — одна из важнейших функций организма. Прекращение дыхания влечет за собой неминуемую гибель человека уже через 3-5 мин. Запасы кислорода в организме очень незначительны, поэтому необходимо его постоянное поступление через систему внешнего дыхания. Указанное обстоятельство объясняет формирование в процессе эволюции такого механизма регуляции, который должен обеспечивать высокую надежность выполнения дыхательных движений. В основе деятельности системы регуляции дыхания лежит поддержание константного уровня таких показателей организма, как Рсо2, Ро2, рН. Их содержание остается относительно постоянным , несмотря на различные потребности в нем, которые во время интенсивной мышечной работы могут увеличиваться в 20 раз.
Регуляция дыхания направлена на выполнение двух задач: во-первых, автоматическая генерация частоты и силы сокращения дыхательных мышц, во-вторых, подстройка ритма и глубины дыхательных движений к реальным потребностям организма (в первую очередь, к изменениям метаболических параметров в виде рo2, рco2 и pH артериальной крови и рco2 и рH межклеточной жидкости мозга).

Рис. 20. Нервный контроль вентиляции
Дыхание регулируется рефлекторным путем, включающим три составных элемента (рис. 20):
-рецепторы, воспринимающие информацию, и афферентные пути, передающие ее нервным центрам;
— нервные центры;
— эффекторы – пути передачи команд от центров и сами регулируемые объекты.
На рисунке 20 представлены образующие систему регуляции дыхания блоки — контролирующие (нервные центры), исполнительные (дыхательные мышцы), рецепторные (хемо- и механорецепторы) и связи между этими блоками. Таким образом, вся система регуляции дыхания состоит из нескольких взаимосвязанных регуляторных контуров.
Входящие сигналы. Генератор ритма получает нисходящую из коры головного мозга импульсацию, а также нервные сигналы от нервных клеток интегратора сенсорной информации и непосредственно от центральных хеморецепторов.
Выходящие сигналы. Нервная импульсация от генератора ритма направляется к иннервирующим дыхательные мышцы двигательным нервным клеткам соответствующих ядер черепных нервов (VII, IX–XII) и к мотонейронам передних рогов спинного мозга (их аксоны в составе спинномозговых нервов направляются к дыхательным мышцам).
Интегратор сенсорной информации получает чувствительную информацию от разнообразных хемо- и механорецепторов, расположенных в органах дыхания и дыхательных мышцах, по ходу магистральных кровеносных сосудов (периферические хеморецепторы), а также в продолговатом мозге (центральные хеморецепторы). Помимо этих прямых сигналов, интегратор получает множество информации, опосредованной различными структурами мозга (в том числе и от высших отделов ЦНС). Импульсация от нервных клеток интегратора, направляясь к нейронам генератора ритма, модулирует характер разрядов от них.
Контрольные вопросы
1. Что лежит в основе деятельности системы регуляции дыхания ?
2. Какие составные элементы образуют систему регуляции дыхания ?
3. Какова роль интеграции сенсорной информации в системе регуляции дыхания ?
Дыхательный центр
Дыхательный центр — совокупность нейронных ансамблей разных этажей центральной нервной системы, обеспечивающих управление внешним дыханием (рис. 21).

Рис. 21. Уровни организации дыхательного центра
Главная часть дыхательного центра находится в продолговатом мозге. Это было установлено в опытах с перерезкой ствола мозга, проведенных французским физиологом Легуллуа в 1812 г. Миславский Н.А. (отечественный ученый) установил наличие двух структур, ответственных за вдох и выдох, которые расположены в продолговатом мозге.
Автоматический дыхательный центр — совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм (генераторы ритма).
Основная часть нейронов дыхательного центра продолговатого мозга относится к ретикулярной формации, эти нейроны обладают свойством спонтанной активности.
Дыхательный центр выполняет две основные функции в системе дыхания:
1) моторную, или двигательную, которая проявляется в виде сокращения дыхательных мышц
2) гомеостатическую, связанную с изменением характера дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна (длительность вдоха и выдоха). Моторная функция дыхательного центра адаптирует дыхание к метаболическим потребностям организма, приспосабливает дыхание в поведенческих реакциях (поза, бег и др.), а также осуществляет интеграцию дыхания с другими функциями ЦНС.
Гомеостатическая функция дыхательного центра поддерживает нормальные величины дыхательных газов (O2, CO2) и рН в крови и внеклеточной жидкости мозга, регулирует дыхание при изменении температуры тела, адаптирует дыхательную функцию к условиям измененной газовой среды, например при пониженном и повышенном барометрическом давлении.
Нейроны дыхательного центра локализованы в дорсомедиальной и вентролатеральной областях продолговатого мозга и образуют так называемые дорсальную и вентральную дыхательную группу (рис. 22).
Дыхательные нейроны, активность которых вызывает инспирацию или экспирацию, называются соответственно инспираторными и экспираторными нейронами. Инспираторные и экспираторные нейроны иннервируют дыхательные мышцы.

Рис. 22. Проекция местоположения дыхательного центра на дорсальную поверхность продолговатого мозга. ДДГ и ВДГ — соответственно дорсальная и вентральная дыхательные группы; Бк — комплекс Бетцингера; рВДГ и кВДГ — ростральная и каудальная часть ВДГ; СI—СII — сегменты спинного мозга; ДН, НМ и ВМ — соответственно диафрагмальный нерв и нервы наружных и внутренних межреберных мышц.
Дорсальная дыхательная группа (ДДГ) включает в себя симметричные области продолговатого мозга, расположенные вентролатеральнее ядра одиночного пучка (рис. 22).
Большинство нейронов этой группы (около 90 %) являются инспираторными. Нейроны ДДГ получают афферентные сигналы от легочных рецепторов растяжения по волокнам блуждающего нерва, нейроны которого имеют обширные синаптические связи с другими отделами дыхательного центра и с различными отделами ЦНС. Только часть инспираторных нейронов ДДГ связана аксонами с дыхательными мотонейронами спинного мозга, преимущественно с контралатеральной стороны. Другая часть нейронов взаимодействует с нейронами вентральной группы. Базовый ритм дыхания генерируется в основном дорсальной группой дыхательных нейронов. Даже после перерезки всех входящих в продолговатый мозг периферических нервов и ствола мозга выше и ниже продолговатого мозга эта группа нейронов продолжает генерировать потенциалы действия.
Вентральная дыхательная группа (ВДГ) расположена латеральнее обоюдного ядра продолговатого мозга, или ядра блуждающего нерва. ВДГ подразделяется на ростральную и каудальную части относительно уровня задвижки (obex) продолговатого мозга (рис. 22). Нейроны вентральной группы подразделяются на две части:
1) Ростральная часть ВДГ состоит из инспираторных нейронов, часть которых взаимодействует с другими дыхательными нейронами продолговатого мозга, другая часть нейронов направляют свои аксоны к дыхательным мотонейронам спинного мозга, а следовательно, управляют мышцами вдоха.
2) Каудальная часть ВДГ состоит только из экспираторных нейронов. Все экспираторные нейроны направляют аксоны в спинной мозг. При этом 40% экспираторных нейронов иннервирует внутренние межреберные мышцы, а 60% — мышцы брюшной стенки.
Функции нейронов вентральной группы отличаются от функций нейронов дорсальной группы:
а) во время спокойного дыхания нейроны вентральной группы остаются почти полностью неактивными (выдох совершается под влиянием эластической тяги легких и грудной клетки);
б) нейроны не участвуют в основной ритмической осцилляции, регулирующей дыхание;
в) при форсированном дыхании нейроны вентральной группы подключаются к нейронам дорсальной группы и участвуют в формировании выдоха (усиливают выдох при его затруднении и тяжелой мышечной нагрузке).
Ростральнее ВДГ локализованы компактной группой экспираторные нейроны (комплекс Бетцингера), аксоны которых связаны только с другими типами нейронов дыхательного центра. Предполагают, что именно эти нейроны синхронизируют деятельность правой и левой половин дыхательного центра.
В связи с тем, что дыхательный цикл состоит из вдоха и выдоха, имеется несколько классификаций нейронов дыхательного центра.

Рис. 23. Активность основных групп дыхательных нейронов: 1 — ранние; 2 — полные; 3 — поздние инспираторные; 4 — постинспираторные; 5 – экспираторные; 6 — преинспираторные нейроны
Выделяют следующие основные типы дыхательных нейронов: 1) ранние инспираторные, которые разряжаются с максимальной частотой в начале фазы вдоха; 2) поздние инспираторные, максимальная частота разрядов которых приходится на конец инспирации; 3) полные инспираторные с постоянной или с постепенно нарастающей активностью в течение фазы вдоха; 4) постинспираторные, которые имеют максимальный разряд в начале фазы выдоха; 5) экспираторные с постоянной или постепенно нарастающей активностью, которую они проявляют во вторую часть фазы выдоха; 6) преинспираторные, которые имеют максимальный пик активности непосредственно перед началом вдоха. Имеются и другие классификации дыхательных нейронов. Тип нейронов определяется по проявлению его активности относительно фазы вдоха и выдоха (рис. 23).
Нейроны дыхательного центра, активность которых совпадает с ритмом дыхания, но они не иннервируют дыхательные мышцы, называются респираторно-связанными нейронами. К респираторно-связанным нейронам относят клетки дыхательного центра, иннервирующие мышцы верхних дыхательных путей, например гортани.
Нейроны дыхательного центра в зависимости от проекции их аксонов подразделяют на три группы: 1) нейроны, иннервирующие мышцы верхних дыхательных путей и регулирующие поток воздуха в дыхательных путях; 2) нейроны, которые синаптически связаны с дыхательными мотонейронами спинного мозга и управляют таким образом мышцами вдоха и выдоха; 3) проприобульбарные нейроны, которые связаны с другими нейронами дыхательного центра и участвуют только в генерации дыхательного ритма.
Контрольные вопросы
1. Какие отделы мозга входят в структуру дыхательного центра ?
2. Что такое автоматический дыхательный центр, где он расположен ?
3. Какие основные функции выполняет дыхательный центр ?
4. Какие нейроны входят в дорсальную дыхательную группу, их функции ?
5. Какие нейроны входят в вентральную дыхательную группу, их функции ?
6. Какова роль комплекса Бетцингера в регуляции дыхания ?
7. Какие существуют основные типы дыхательных нейронов ?
6.2. Другие области локализации дыхательных нейронов
В мосту находятся два ядра дыхательных нейронов: медиальное парабрахиальное ядро и ядро Шатра (ядро Келликера). Иногда эти ядра называют пневмотаксическим центром. Главным в деятельности этого центра является контроль за точкой «выключения» нарастающего инспираторного сигнала и длительностью фазы вдоха. Первичной задачей пневмотаксического центра является укорочение вдоха. При этом возникает вторичный эффект – увеличение скорости дыхания, т.к. ограничение вдоха укорачивает длительность выдоха. Это может увеличивать частоту дыхания до 30-40 в минуту.
Некоторые авторы полагают, что в нижних отделах варолиева моста расположен еще один — апнейстический центр. Если произвести перерезку варолиева моста и отделить пневмотаксический центр, оставив бульбарный и апнейстический, то у животных можно наблюдать продолжительные судорожные вдохи, прерываемые короткими выдохами.
Диафрагмальные мотонейроны. Образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Диафрагмальный нерв состоит из 700—800 миелинизированных и более 1500 немиелинизированных волокон. Подавляющее количество волокон является аксонами б-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен, локализованных в диафрагме, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны сегментов спинного мозга, иннервирующие дыхательные мышцы. На уровне CI—СII вблизи латерального края промежуточной зоны серого вещества находятся инспираторные нейроны, которые участвуют в регуляции активности межреберных и диафрагмальных мотонейронов. Мотонейроны, иннервирующие межреберные мышцы, локализованы в сером веществе передних рогов на уровне от TIV до ТX. Причем одни нейроны регулируют преимущественно дыхательную, а другие — преимущественно позно-тоническую активность межреберных мышц. Мотонейроны, иннервирующие мышцы брюшной стенки, локализованы в пределах вентральных рогов спинного мозга на уровне TIV—LIII.
Контрольные вопросы
1. Что такое пневмотаксический центр, где он расположен ?
2. Что такое апнейстический центр, где он расположен ?
3. Какова роль мотонейронов спинного мозга в регуляции дыхания ?
§
Ритмическое сокращение и расслабление дыхательных мышц обеспечивается циркуляцией возбуждения в дыхательных нейронах продолговатого мозга, а также взаимодействием импульсации нейронов продолговатого мозга с импульсацией дыхательных нейронов моста. ____________________________________________________________________________________________________________Дыхательный цикл подразделяют на фазу вдоха и фазу выдоха относительно движения воздуха из атмосферы в сторону альвеол (вдох) и обратно (выдох). Двум фазам внешнего дыхания соответствуют три фазы активности нейронов дыхательного центра продолговатого мозга: инспираторная, которая соответствует вдоху; постинспираторная, которая соответствует первой половине выдоха и называется пассивной контролируемой экспирацией; экспираторная, которая соответствует второй половине фазы выдоха и называется фазой активной экспирации (рис. 24).
Дыхательный цикл начинается с возбуждения ранних инспираторных нейронов, которые полностью освобождаются от сильного торможения со стороны постинспираторных нейронов. Полное растормаживание ранних инспираторных нейронов происходит в момент, когда активируются преинспираторные нейроны дыхательного центра, которые окончательно блокируют разряд экспираторных нейронов. Далее возбуждение переходит на полные инспираторные нейроны, которые способны совозбуждать друг друга. Полные инспираторные нейроны, благодаря этому свойству, поддерживают и увеличивают частоту генерации потенциалов действия в течение фазы вдоха. Именно этот тип дыхательных нейронов создает нарастающую активность в диафрагмальном и межреберных нервах, вызывая увеличение силы сокращения диафрагмы и наружных межреберных мышц.

Рис. 24. Соотношение фаз дыхательного цикла и фаз активности нейронов дыхательного центра
Ранние инспираторные нейроны в силу особых физиологических свойств их мембраны прекращают генерировать потенциалы действия к середине фазы вдоха. Это моносинаптически растормаживает поздние инспираторные нейроны, поэтому их активность появляется в конце вдоха.
Поздние инспираторные нейроны способны дополнительно активировать в конце вдоха сокращение диафрагмы и наружных межреберных мышц. Одновременно поздние инспираторные нейроны выполняют функцию начального выключения инспирации. В период своей активности они получают возбуждающие стимулы от легочных рецепторов растяжения, которые измеряют степень растяжения дыхательных путей во время вдоха. Максимальный по частоте разряд поздних инспираторных нейронов приходится на момент прекращения активности других типов инспираторных нейронов дыхательного центра.
Прекращение активности всех типов инспираторных нейронов дыхательного центра растормаживает постинспираторные нейроны. Причем процесс растормаживания постинспираторных нейронов начинается гораздо раньше, а именно в период убывания разрядов ранних инспираторных нейронов. С момента появления активности постинспираторных нейронов выключается инспирация и начинается фаза пассивной контролируемой экспирации. Постинспираторные нейроны регулируют степень расслабления диафрагмы в первую половину фазы выдоха. В эту фазу заторможены все другие типы нейронов дыхательного центра. Однако в постинспираторную фазу сохраняется активность респираторно-связанных нейронов дыхательного центра, которые регулируют тонус мышц верхних дыхательных путей, прежде всего гортани.
Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от механизма ритмогенеза инспираторной и постинспираторной активности. Например, при быстрых дыхательных движениях постинспираторная фаза может непосредственно переходить в фазу следующей инспирации.
Активность дыхательных мышц в течение трех фаз нейронной активности дыхательного центра тоже изменяется (рис. 24). В инспирацию мышечные волокна диафрагмы и наружных межреберных мышц постепенно увеличивают силу сокращения. В этот же период активируются мышцы гортани, которые расширяют голосовую щель, что снижает сопротивление воздушному потоку на вдохе. Работа инспираторных мышц во время вдоха создает достаточный запас энергии, которая высвобождается в постинспираторную фазу, или в фазу пассивной контролируемой экспирации. В постинспираторную фазу дыхания объем выдыхаемого из легких воздуха контролируется медленным расслаблением диафрагмы и одновременным сокращением мышц гортани. Сужение голосовой щели в постинспираторную фазу увеличивает сопротивление воздушному потоку на выдохе. Это является очень важным физиологическим механизмом, который препятствует спадению воздухоносных путей легких при резком увеличении скорости воздушного потока на выдохе, например при форсированном дыхании или защитных рефлексах кашля и чиханья.
Во вторую фазу выдоха, или фазу активной экспирации, экспираторный поток воздуха усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки. В эту фазу отсутствует электрическая активность диафрагмы и наружных межреберных мышц.
Координация деятельности правой и левой половин дыхательного центра является еще одной функцией дыхательных нейронов. Дыхательный центр имеет дорсальную и вентральную группу нейронов как в правой, так и в левой половине продолговатого мозга и таким образом состоит из двух симметричных половин. Эта функция выполняется за счет синаптического взаимодействия различных типов дыхательных нейронов. Дыхательные нейроны взаимосвязаны как в пределах одной половины дыхательного центра, так и с нейронами противоположной стороны. При этом наибольшее значение в синхронизации деятельности правой и левой половин дыхательного центра имеют проприобульбарные дыхательные нейроны и экспираторные нейроны комплекса Бетцингера.
Таким образом, смена вдоха на выдох обеспечивается торможением ранних инспираторных нейронов за счет трех источников импульсов: 1) от нейронов моста; 2) от рецепторов легких, импульсация от которых поступает по блуждающим нервам; 3) от самих инспираторных нейронов.
Контрольные вопросы
1. Какие существуют фазы активности нейронов дыхательного центра продолговатого мозга ?
2. С возбуждения каких нейронов начинается дыхательный цикл ?
3. Какие нейроны активны в инспираторную фазу ?
4. Какие нейроны активны в постинспираторную фазу ?
5. Какие нейроны активны в экспираторную фазу ?
6. Как изменяется активность дыхательных мышц в течение трех фаз нейронной активности дыхательного центра ?
7. Какие нейроны играют важную роль в синхронизации деятельности правой и левой половин дыхательного центра ?
§
| Высота над уровнем моря, м | Атмосферное давление, мм рт. ст. | Парциальное давление кислорода, мм рт. ст |
Снижение парциального давления кислорода во вдыхаемом воздухе приводит к еще более низкому уровню его в альвеолах и оттекающей крови. Если жители равнин поднимаются в горы, гипоксия увеличивает у них вентиляцию легких, стимулируя артериальные хеморецепторы. Организм реагирует приспособительными реакциями, цель которых – улучшение обеспечения тканей О2. Изменения дыхания при высотной гипоксии у разных людей различны. Возникающие во всех случаях реакции внешнего дыхания определяются рядом факторов: 1) скорость, с которой развивается гипоксия; 2) степень потребления О2 (покой или физическая нагрузка); 3) продолжительность гипоксического воздействия.
Важнейшей компенсаторной реакцией на гипоксию является гипервентиляция. Первоначальная гипоксическая стимуляция дыхания, возникающая при подъеме на высоту, приводит к вымыванию из крови СО2 и развитию дыхательного алкалоза. Это в свою очередь вызывает увеличение рН внеклеточной жидкости мозга. Центральные хеморецепторы реагируют на подобный сдвиг рН в цереброспинальной жидкости мозга резким снижением своей активности, что затормаживает нейроны дыхательного центра настолько, что он становится нечувствительным к стимулам, исходящим от периферических хеморецепторов. Довольно быстро гиперпноэ сменяется непроизвольной гиповентиляцией, несмотря на сохраняющуюся гипоксемию. Подобное снижение функции дыхательного центра увеличивает степень гипоксического состояния организма, что чрезвычайно опасно, прежде всего для нейронов коры большого мозга.
При акклиматизации к условиям высокогорья наступает адаптация физиологических механизмов к гипоксии. После пребывания в течение от нескольких дней или недель на высоте, как правило респираторный алкалоз компенсируется за счет выделения почками НСО3, благодаря чему часть тормозного влияния на альвеолярную гипервентиляцию выпадает и гипервентиляция усиливается. Акклиматизация вызывает, кроме того рост концентрации гемоглобина вследствие повышения гипоксической стимуляции почками эритропоэтинов. Так, у жителей Анд, постоянно живущих на высоте 5000 м, концентрация гемоглобина в крови составляет 200 г/л. Основными средствами адаптации к гипоксии являются: 1) значительное увеличение легочной вентиляции; 2) увеличение количества эритроцитов; 3) увеличение диффузионной способности легких; 4) увеличение васкуляризации периферических тканей; 5) увеличение способности клеток тканей использовать кислород, несмотря на низкий рО2.
У некоторых людей при быстром подъеме на большую высоту развивается острое патологическое состояние (острая горная болезнь и высотный отек легких). Так как из всех органов ЦНС обладает высочайшей чувствительностью к гипоксии, то при подъеме на большие высоты в первую очередь возникают неврологические нарушения. При подъеме на высоту могут остро развиться такие симптомы, как головная боль, усталость, тошнота. Часто наступает отек легких. Ниже 4500 м подобные тяжелые нарушения наступают реже, хотя возникают небольшие функциональные отклонения. В зависимости от индивидуальных особенностей организма и его способности к акклиматизации человек способен достигать большой высоты.
Контрольные вопросы
1. Как изменяются параметры барометрического давления и парциального давления кислорода с увеличением высоты над уровнем моря ?
2. Какие приспособительные реакции возникают при подъеме на высоту ?
3. Как происходит акклиматизация к условиям высокогорья ?
4. Как проявляется острая горная болезнь ?
Дыхание при погружении на глубину
При производстве подводных работ водолаз дышит под давлением выше атмосферного на 1 атм. на каждые 10 м погружения. Около 4/5 воздуха составляет азот. При давлении на уровне моря азот не оказывает на организм никакого существенного влияния, но при высоком давлении он может вызывать различные степени наркоза. Первые признаки умеренного наркоза появляются на глубине около 37 м, если водолаз остается на глубине в течение часа или больше и дышит сжатым воздухом. При длительном пребывании на глубине более 76 м (давление 8,5 атм.) обычно развивается азотный наркоз, проявления которого похожи на алкогольную интоксикацию. Если человек вдыхает воздух обычного состава, то происходит растворение азота в жировой ткани. Диффузия азота из тканей происходит медленно, поэтому подъем водолаза на поверхность должен осуществляться очень медленно. В противном случае возможно внутрисосудистое образование пузырьков азота (кровь «закипает») с тяжелыми повреждениями ЦНС, органов зрения, слуха, сильными болями в области суставов. Возникает так называемая кессонная болезнь. Для лечения пострадавшего необходимо вновь поместить в среду с высоким давлением. Постепенная декомпрессия может продолжаться несколько часов или суток.
Вероятность возникновения кессонной болезни может быть значительно снижена при дыхании специальными газовыми смесями, например кислородно-гелиевой смесью. Это связано с тем, что растворимость гелия меньше, чем азота, и он быстрее диффундирует из тканей, так как его молекулярная масса в 7 раз меньше, чем у азота. Кроме того, эта смесь обладает меньшей плотностью, поэтому уменьшается работа, затрачиваемая на внешнее дыхание.
Контрольные вопросы
5. Как изменяются параметры барометрического давления и парциального давления кислорода с увеличением высоты над уровнем моря ?
6. Какие приспособительные реакции возникают при подъеме на высоту ?
7. Как происходит акклиматизация к условиям высокогорья ?
8. Как проявляется острая горная болезнь ?
7.3 Тестовые задания и ситуационная задача
Выберите один правильный ответ.
41. ЕСЛИ ЧЕЛОВЕК НЫРЯЕТ БЕЗ СПЕЦИАЛЬНОГО СНАРЯЖЕНИЯ С ПРЕДВАРИТЕЛЬНОЙ ГИПЕРВЕНТИЛЯЦИЕЙ, ПРИЧИНОЙ ВНЕЗАПНОЙ ПОТЕРИ СОЗНАНИЯ МОЖЕТ СТАТЬ НАРАСТАЮЩАЯ
1) асфиксия
2) гипоксия
3) гипероксия
4) гиперкапния
42. ПРИ ПОГРУЖЕНИИ ПОД ВОДУ С МАСКОЙ И ТРУБКОЙ НЕЛЬЗЯ УВЕЛИЧИВАТЬ ДЛИНУ СТАНДАРТНОЙ ТРУБКИ (30-35 см) ИЗ-ЗА
1) возникновения градиента давления между давлением воздуха в альвеолах и давлением воды на грудную клетку
2) опасности возникновения гиперкапнии
3) опасности возникновения гипоксии
4) увеличения объёма мёртвого пространства
Ситуационная задача 8
Чемпионы по нырянию погружаются на глубину до 100 м без акваланга и возвращаются на поверхность за 4-5 минут. Почему у них не возникает кессонная болезнь ?
8. Эталоны ответов к тестовым заданиям и ситуационным задачам
Эталоны ответов к тестовым заданиям:
1-3
2-1
3-2
4-3
5-2
6-3
7-3
8-3
9-3
10-4
11-2
12-3
13-1
14-3
15-2
16-1
17-4
18-2
19-3
20-1
21-3
22-1
23-4
24-3
25-2
26-3
27-4
28-2
29-1
30-2
31-2
32-2
33-3
34-4
35-1
36-4
37-4
38-3
39-2
40-4
41-2
42-1
Эталоны ответов к ситуационным задачам:
Решение ситуационной задачи № 1:
Если речь идет об естественном дыхании, то прав первый. Механизм дыхания всасывающий. Но, если иметь в виду искусственное дыхание, то прав второй, так как здесь механизм нагнетательный.
Решение ситуационной задачи № 2:
Для эффективного газообмена необходимо определенное соотношение между вентиляцией и кровотоком в сосудах легких. Следовательно, у этих людей имелись различия в величинах кровотока.
Решение ситуационной задачи № 3:
В крови кислород находится в двух состояниях: физически растворенном и связанном с гемоглобином. Если гемоглобин работает плохо, то остается только растворенный кислород. Но его очень мало. Значит необходимо увеличить его количество. Это достигается путем гипербарической оксигенации (пациента помещают в камеру с высоким давлением кислорода).
Решение ситуационной задачи № 4:
Малат окисляется НАД-зависимым ферментом малатдегидрогеназой (митохондриальной фракцией). Причем при окислении одной молекулы малата образуется одна молекула НАДН·Н , которая вступает в полную цепь переноса электронов с образованием из трех молекул АДФ трех молекул АТФ. Как известно АДФ является активатором дыхательной цепи, а АТФ — ингибитором. АДФ по отношению к малату взято заведомо в недостатке. Это приводит к тому, что из системы исчезает активатор (АДФ) и появляется ингибитор (АТФ), что, в свою очередь, приводит к остановке дыхательной цепи и поглощению кислорода. Гексокиназа катализирует перенос фосфатной группы с АТФ на глюкозу с образованием глюкозо-6-фосфата и АДФ. Таким образом, при работе этого фермента в системе расходуется ингибитор (АТФ) и появляется активатор (АДФ), поэтому дыхательная цепь возобновила работу.
Решение ситуационной задачи № 5:
Фермент сукцинатдегидрогеназа, катализирующий окисление сукцината, относится к ФАД-зависимым дегидрогеназам. Как известно ФАДН2 обеспечивает поступление водорода в укороченную цепь переноса электронов в ходе которой образуется 2 молекулы АТФ. Амобарбитал блокирует дыхательную цепь на уровне 1-го сопряжения дыхания и фосфорилирования и на окисление сукцината не влияет.
Решение ситуационной задачи № 6:
При очень медленном пережатии пуповины соответственно очень медленно будет нарастать содержание углекислого газа в крови и нейроны дыхательного центра не смогут возбудиться. Первый вдох так и не произойдет.
Решение ситуационной задачи № 7:
Ведущую роль в возбуждении нейронов дыхательного центра играет углекислый газ. При агональном состоянии возбудимость нейронов дыхательного центра резко снижается и поэтому они не могут возбуждаться при действии обычных количеств углекислого газа. После нескольких дыхательных циклов наступает пауза, во время которой накапливаются значительные количества углекислого газа. Теперь они уже могут возбудить дыхательный центр. Происходит несколько вдохов-выдохов, количество углекислого газа снижается, снова наступает пауза и т.д. Если не удается улучшить состояние больного, неизбежен летальный исход.
Решение ситуационной задачи № 8:
Водолаз на большой глубине дышит воздухом под высоким давлением. Поэтому растворимость газов в крови значительно возрастает. Азот в организме не потребляется. Поэтому при быстром поднятии его повышенное давление быстро снижается , и он бурно выделяется из крови в виде пузырьков, что приводит к эмболии. Ныряльщик же во время погружения вообще не дышит. При быстром поднятии ничего страшного не происходит.
Приложение 1
Таблица 1
Наименование показателей легочной вентиляции на русском и английском языках
| Наименование показателя на русском языке | Принятое сокращение | Наименование показателя на английском языке | Принятое сокращение |
| Жизненная емкость легких | ЖЕЛ | Vital capacity | VC |
| Дыхательный объем | ДО | Tidal volume | TV |
| Резервный объем вдоха | РОвд | Inspiratory reserve volume | IRV |
| Резервный объем выдоха | РОвыд | Expiratory reserve volume | ERV |
| Максимальная вентиляция легких | МВЛ | Maximal voluntary ventilation | MW |
| Форсированная жизненная емкость легких | ФЖЕЛ | Forced vital capacity | FVC |
| Объем форсированного выдоха за первую секунду | ОФВ1 | Forced expiratory volume 1 sec | FEV1 |
| Индекс Тиффно | ИТ, или ОФВ1/ЖЕЛ % | FEV1 % = FEV1/VC % | |
| Максимальная объемная скорость в момент выдоха 25 % ФЖЕЛ, оставшейся в легких | МОС25 | Maximal expiratory flow 25 % FVC | MEF25 |
| Forced expiratory flow 75 % FVC | FEF75 | ||
| Максимальная объемная скорость в момент выдоха 50 % ФЖЕЛ, оставшейся в легких | МОС50 | Maximal expiratory flow 50 % FVC | MEF50 |
| Forced expiratory flow 50 % FVC | FEF50 | ||
| Максимальная объемная скорость в момент выдоха 75 % ФЖЕЛ, оставшейся в легких | МОС75 | Maximal expiratory flow 75 % FVC | MEF75 |
| Forced expiratory flow 25 % FVC | FEF25 | ||
| Средняя объемная скорость выдоха в интервале от 25 % до 75 % ФЖЕЛ | СОС25-75 | Maximal expiratory flow 25-75 % FVC | MEF25-75 |
| Forced expiratory flow 25-75 % FVC | FEF25-75 |
Приложение 2
ОСНОВНЫЕ ПАРАМЕТРЫ ДЫХАНИЯ
ЖЕЛ (VC = Vital Capacity) — жизненная ёмкость лёгких (объём воздуха, который выходит из лёгких при максимально глубоком выдохе после максимально глубокого вдоха)
РOвд (IRV = inspiratory reserve volume) — резервный объём вдоха (дополнительный воздух) — это тот объём воздуха, который можно вдохнуть при максимальном вдохе после обычного вдоха
РOвыд (ERV = Expiratory Reserve Volume) — резервный объём выдоха (резервный воздух) — это тот объём воздуха, который можно выдохнуть при максимальном выдохе после обычного выдоха
ЕВ (IC = inspiratory capacity) — емкость вдоха — фактическая сумма дыхательного объёма и резервного объёма вдоха (ЕВ = ДО РОвд)
ФОЕЛ (FRC = functional residual capacity) — функциональная остаточная емкость легких. Это объём воздуха в лёгких пациента, находящегося в состоянии покоя, в положении, когда закончен обычный выдох, а голосовая щель открыта. ФОЕЛ представляет собой сумму резервного объёма выдоха и остаточного воздуха (ФОЕЛ = РОвыд ОВ). Данный параметр можно измерить с помощью одного из двух способов: разведения гелия или плетизмографии тела. Спирометрия не позволяет измерить ФОЕЛ, поэтому значение данного параметра требуется ввести вручную.
ОВ (RV = residual volume) — остаточный воздух (другое название — ООЛ, остаточный объём лёгких) — это объём воздуха, который остается в лёгких после максимального выдоха. Остаточный объём нельзя определить с помощью одной спирометрии; это требует дополнительных измерений объёма легких (с помощью метода разведения гелия или плетизмографии тела).
ОЕЛ (TLC = total lung capacity) — общая емкость легких (объём воздуха, находящийся в лёгких после максимально глубокого вдоха). ОЕЛ = ЖЕЛ ОВ
Расход кислорода при резке труб. резка труб кислородом.
Для вычисления себестоимости работ по резке металла используют нормы расхода кислорода. Производится общий расход кислорода в течении рабочей смены и умножают на стоимость. Чтобы определить расход кислорода при работах по резке труб используют нормы расхода кислорода, приведенные в таблице ниже. В таблице приведены значения для одного перереза разного диаметра и толщины труб.
В предыдущей статье указан расход кислорода для листового металла, а в этой статье, остановимся на нормах расхода кислорода при резке труб. Данные нормы рассчитаны для керосинорезов типа «Бобуха».
Таблица 1.
Расход кислорода для перереза трубы.
| Труба (наружный диаметр× толщина стенки), мм | Расход кислорода, м 3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 14 × 2,0 | 0,00348 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 16 × 3,5 | 0,00564 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 20 × 2,5 | 0,00566 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 32 × 3,0 | 0,0102 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 45 × 3,0 | 0,0143 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 57 × 6,0 | 0,0344 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 76 × 8,0 | 0,0377 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 89 × 6,0 | 0,0473 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 108 × 6,0 | 0,0574 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 114 × 6,0 | 0,0605 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 133 × 6,0 | 0,0705 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 159 × 8,0 | 0,119 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 219 × 12,0 | 0,213 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 426 × 10,0 | 0,351 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ø 530 × 10,0 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Толщина стенки, мм | Расход материалов по видам резки, л, с использованием | Код строки | |||||
| Ацетилена | пропан-бутановой смеси | природного газа | |||||
| Ацетилен | Кислород | Пропан-бутан | Кислород | Природный газ | Кислород | ||
| 3 | 11,98 | 53,92 | 8,72 | 69,08 | 19,49 | 69,08 | 01 |
| 4 | 15,93 | 71,85 | 11,57 | 92,04 | 25,94 | 92,04 | 02 |
| 5 | 19,96 | 89,84 | 14,49 | 115,05 | 32,43 | 115,05 | 03 |
| 6 | 23,95 | 107,81 | 17,39 | 138,03 | 38,93 | 138,03 | 04 |
| 8 | 27,92 | 143,69 | 22,26 | 184 | 49,68 | 184 | 05 |
| 10 | 28,07 | 180,77 | 23 | 230,08 | 50,62 | 230,08 | 06 |
| 12 | 33,62 | 215,55 | 25,66 | 275,98 | 55,95 | 275,98 | 07 |
| 15 | 45,94 | 294,66 | 35,08 | 377,29 | 79,23 | 377,29 | 08 |
| 18 | 46,37 | 335,33 | 36 | 413,99 | 79,69 | 413,99 | 09 |
| 20 | 51,52 | 372,6 | 36,1 | 460 | 81,88 | 460 | 10 |
| 25 | 64,39 | 465,75 | 44,85 | 575 | 102,35 | 575 | 11 |
| Код графы | 01 | 02 | 03 | 04 | 05 | 06 | |
Нормы на 1 перерез Таблица 037
| Толщина стенки, мм | Расход материалов по видам резки, л, с использованием | Код строки | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ацетилена | пропан-бутановой смеси | природного газа | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ацетилен | Кислород | Пропан-бутан | Кислород | Природный газ | Кислород | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 45 ´ 3 | 1,58 | 7,11 | 1,15 | 9,11 | 2,57 | 9,11 | 01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 45 ´ 4 | 2,05 | 9,25 | 1,49 | 11,85 | 3,34 | 11,85 | 02 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 57 ´ 3 | 2,03 | 9,14 | 1,48 | 11,71 | 3,3 | 11,71 | 03 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 57 ´ 4 | 2,66 | 11,96 | 1,93 | 15,32 | 4,32 | 15,32 | 04 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 76 ´ 5 | 4,45 | 20,03 | 3,23 | 25,65 | 7,23 | 25,65 | 05 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 89 ´ 5 | 5,26 | 23,69 | 3,82 | 30,34 | 8,55 | 30,34 | 06 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 108 ´ 6 | 7,67 | 34,53 | 5,57 | 44,21 | 12,47 | 44,21 | 07 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 133 ´ 6 | 9,55 | 42,97 | 6,93 | 55,04 | 15,52 | 55,04 | 08 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 133 ´ 8 | 12,53 | 56,4 | 9,09 | 72,22 | 20,37 | 72,22 | 09 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 159 ´ 8 | 15,14 | 68,13 | 10,99 | 87,24 | 24,6 | 87,24 | 10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 219 ´ 6 | 16,01 | 72,1 | 11,63 | 92,33 | 26,03 | 92,33 | 11 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 219 ´ 8 | 18 | 91,46 | 14,75 | 121,95 | 32,92 | 121,95 | 12 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 219 ´ 10 | 18,42 | 118,63 | 15,1 | 150,99 | 33,22 | 150,99 | 13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 219 ´ 12 | 21,86 | 140,99 | 16,69 | 179,53 | 37,69 | 179,53 | 14 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 273 ´ 8 | 23 | 119,58 | 18,52 | 153,11 | 41,34 | 153,11 | 15 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 273 ´ 10 | 23,17 | 148,33 | 18,99 | 189,93 | 41,79 | 189,93 | 16 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 273 ´ 12 | 27,55 | 176,65 | 21,03 | 226,18 | 47,5 | 226,18 | 17 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 273 ´ 15 | 37,22 | 238,71 | 28,42 | 305,65 | 64,19 | 305,65 | 18 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 325 ´ 8 | 27 | 143,04 | 22,16 | 183,15 | 49,45 | 183,15 | 19 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 325 ´ 10 | 27,75 | 177,67 | 22,75 | 227,49 | 50,05 | 227,49 | 20 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 325 ´ 12 | 33,04 | 211,85 | 25,23 | 271,26 | 56,96 | 271,26 | 21 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 325 ´ 15 | 40,9 | 262,27 | 31,23 | 335,82 | 70,52 | 335,82 | 22 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 377 ´ 8 | 31,99 | 166,5 | 25,79 | 213,18 | 57,55 | 213,18 | 23 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 377 ´ 10 | 32,34 | 207 | 26,5 | 265,05 | 58,31 | 265,05 | 24 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 377 ´ 12 | 38,53 | 247,04 | 29,42 | 316,32 | 66,43 | 316,32 | 25 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 377 ´ 15 | 47,76 | 306,27 | 36,47 | 392,15 | 82,35 | 392,15 | 26 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 426 ´ 10 | 36,59 | 234,63 | 27,94 | 300,43 | 63,09 | 300,43 | 27 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 426 ´ 12 | 43,69 | 280,21 | 33,36 | 358,78 | 75,34 | 358,78 | 28 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 426 ´ 15 | 54,23 | 347,72 | 41,41 | 445,23 | 93,5 | 445,23 | 29 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 8 | 39,81 | 206,21 | 31,94 | 264,04 | 71,29 | 264,04 | 30 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 10 | 40,11 | 256,64 | 32,86 | 328,6 | 72,29 | 328,6 | 31 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 12 | 47,82 | 306,61 | 36,51 | 392,59 | 82,44 | 392,59 | 32 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 15 | 59,38 | 380,73 | 45,34 | 487,49 | 102,37 | 487,49 | 33 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 18 | 65,08 | 470,67 | 46,32 | 581,07 | 103,43 | 581,07 | 34 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 20 | 71,99 | 520,63 | 50,18 | 642,76 | 114,41 | 642,76 | 35 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 465 ´ 25 | 88,97 | 643,48 | 61,96 | 794,42 | 141,41 | 794,42 | 36 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 530 ´ 6 | 39,39 | 177,33 | 28,61 | 227,06 | 64,03 | 227,06 | 37 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 530 ´ 8 | 45,33 | 235,55 | 36,49 | 301,6 | 81,43 | 301,6 | 38 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 530 ´ 10 | 45,82 | 293,29 | 37,55 | 375,54 | 82,62 | 375,54 | 39 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 530 ´ 12 | 54,68 | 350,6 | 41,75 | 448,91 | 94,27 | 448,91 | 40 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 530 ´ 15 | 67,95 | 435,73 | 51,88 | 557,91 | 117,16 | 557,91 | 41 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 630 ´ 6 | 46,91 | 211,17 | 34,07 | 270,39 | 76,25 | 270,39 | 42 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 630 ´ 8 | 54,35 | 280,67 | 43,49 | 359,38 | 97,03 | 359,38 | 43 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 630 ´ 10 | 54,63 | 349,7 | 44,77 | 447,76 | 98,51 | 447,76 | 44 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 630 ´ 12 | 65,23 | 418,29 | 49,81 | 535,58 | 112,47 | 535,58 | 45 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 630 ´ 15 | 81,15 | 520,33 | 61,96 | 666,24 | 139,91 | 666,24 | 46 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Код графы | 01 | 02 | 03 | 04 | 05 | 06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Примечание. При резке со скосом кромок под углом 50° и 30° нормы необходимо увеличивать соответственно в 1,55 и 1,16 раза. Сколько баллонов кислорода нужно на резку данного объема металла? Данный вопрос является основополагающим, как при вычислении общих затрат в течение трудового процесса, так и при вычислении себестоимости изготовления детали и производства определённых видов работ. Так как кислород является топливом для резки детали, то норма расхода кислорода на резку металла приобретает ключевое значение, наряду с расходом электроэнергии. Существует несколько способов термического разделения металлов, которые подразделяются в зависимости от способа и вида используемого топлива. Поэтому наряду с кислородной резкой металлов мы в данной статье обратим внимание и на другие способы резки металлических конструкций. Итак, приступим. Разновидности термической резки металла. Рассмотрим три основных способа терморезки. Первый по распространению тип – это кислородно-автогенная резка. Область применения – раскрой листового и сортового углеродистого, низколегированного металлопроката, обрезка лишних выступов и кромок, которые образовались во время литья, подготовка деталей под сварку, разделка металлолома и прочее. Данный способ не применяется для разделения нержавеющих высоколегированных сталей, цветных металлов и чугуна. Следующий тип – это плазменно-дуговая резка. Область применения – это также раскрой, но в данном случае низко- и высоколегированных сталей, а также алюминия, меди и их сплавов. И последний тип, который мы рассмотрим в данной статье – это лазерная резка, которая является одним из инновационных методов резки металлов. Этот способ значительно расширяет область применения газовой резки и, благодаря этому, можно эффективно разделять тонколистный прокат, специальный профильный прокат, тонкостенные трубы, как из металлических, так и не из металлических деталей. Расход газового топлива в различных способах (кислород, ацетилен, пропан) на разделение определяется по специализированным таблицам в зависимости от режима резки, а также от толщины разрезаемого металла. При вышеупомянутых типах резки по видам топлива номинируется расход газов, которые используются для разогрева разрезаемой конструкции, для резки, а также для образования плазмы. Повторим, что к таким относятся: кислород, газы-заменители (пропан- бутан, природный газ и др.), ацетилен, а также азот. Кроме этих газов, используются водород и аргон, но их область и популярность применения не значительна, поэтому включать в содержание статьи мы их не будем. Во время работы с плазменно-дуговым прибором важно заранее планировать количество сменных специальных электродов (катодов), с циркониевыми или гафниевыми вставками. Нормы расхода данных электродов меняются в зависимости от интенсивности рабочего процесса и в общем, не превышают 4 стержней за одну смену. Более точное нормирование расхода стержней будет указано в инструкции по эксплуатации данного агрегата. Расход газов на резку металла: нормы. Расхода кислорода на резку металла, как и расход других газов, рассчитывается по специальной формуле: И в этом уравнении Н – это нормативы расхода во время рабочего процесса, кубический метр газа на метр реза. L – величина разреза или вырезаемой детали, метр. Kh – это коэффициент, который учитывает множество особенностей рабочего процесса: расход газа на начальном этапе резке, продувка и регулировка, зажигание плазменной дуги, на прогрев металла, и, как правило, он равняется 1.1 при единичном производстве, или 1.05 — при промышленном производстве. Норма расхода кислорода на резку металла и прочих газов (Н, кубический метр на один метр разреза) во время разделения в зависимости от мощностей оборудования и режима резки, высчитывается по следующей формуле: Где Р – это допустимый расход газов, который указан в технических характеристиках используемого оборудования, метр кубический на час, а V – это скорость разделения метр на час. Основные значения газового расхода по различным диапазонам скорости резки для некоторых типов оборудования, которые можно применять для расчётов крупного масштаба в промышленном производстве, приведены в следующей таблице.
Применение различных газов в качестве топлива для разделительных машин по металлу обусловлено требованиями к применяемому оборудованию и в зависимости от технологического процесса. Зависимость толщины и скорости резки от допустимого расхода газа является прямо пропорциональной и данное значение можно легко и просто определить интерполированием. И поэтому можно укрупнено, оценочно совершить вычисление расхода различных газов независимо от вида термической резки металлов, исходя из размеров разреза, толщины металла, а также мощности оборудования. В конце важно отметить, что нормирование расхода газов – это одна из важных особенностей, от которых зависит эффективность и скорость рабочего процесса, поэтому не стоит забывать про нормативы расхода, особенно во время разделения металла в огромных масштабах. Газовая резка металла — классический метод термической резки. Разделение металла осуществляется режущим газом. Это недорогая и экономичная технология, возможна резка листового материала довольно большой толщины, но газовая резка тонких листов весьма проблематична. Следует также учитывать, что воздействие тепла режущим газом очень велико, и охватывает большую зону, что может вызвать термическую усадку. Еще один существенный недостаток газовой резки — у изделий остаются оплавленные края, что не всегда допустимо при монтаже изделий. Чтобы устранить этот дефект с помощью шлифовальной машинки производят дополнительную обработку — удаление наплывов и заусениц, выравнивание поверхности кромки. Поэтому потери материала при газовой резке могут быть около 2 см. Абразивная резка металла осуществляется на стационарном оборудовании — углошлифовальной машине, которую также называют болгаркой. Применяется, как правило, при обработке небольших партий тонкостенного металлопроката, прутков небольших диаметров, арматуры. Преимущества этого метода: кромка без наплывов и смятия, отсутствие механической и термической деформации металла, при резке не происходит нагревание материала, точность реза достигает ± 2 мм. Кроме того, технология абразивной резки является достаточно экологичной за счет применяемого абразива и отсутствия пыли в процессе резки.
Высокоточная ленточнопильная резка применяется для изделий из цветных металлов, легированной и нержавеющей стали, чугуна. Ленточнопильные станки обеспечивают непрерывную резку труб и сортового металлопроката на заготовки заданного размера. Главный инструмент в станках для резки металла — ленточная пила, которая изготавливается из очень прочной быстрорежущей стали, армированная кобальтом или твердым сплавом. Ленточная пила представляет собой замкнутую ленту, на которой расположены режущие зубья. Чаще всего резка металлопроката на ленточнопильных станках происходит перпендикулярно оси оборудования, но есть возможность резать металл и под заранее заданным углом (отличным от 90º), для этого станки оснащены поворотной рамой. Ленточнопильная резка обеспечивает точность реза ± 1-1,5 мм, поверхность реза получается ровная, без заусенцев. С помощью ленточнопильного оборудования можно разрезать большие в сечении заготовки. Поперечная резка сортового проката увеличенного сечения — операция как раз для ленточнопильного станка. При необходимости на этом оборудовании выполнима поперечная резка труб. Прямолинейность реза при этом просто великолепная (ленточная пила не «скашивает» рез). Источник | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
